#paleo fauna
Text





some cenozoic beasties + closeups :]
#cenozoic#cenozoology#cenozoic era#cenozoic fauna#paleoart#paleo#paleolithic#paleo art#paleo drawing#eohippus#phenacodus#coryphodon#dinocrocuta#phenacodontidae#prehistoric horse#prehistoric animals#cenozoic art#cenozic illustration#cenozoic animals#art#illustration#artists on tumblr#animal art#animal illustration#comics#doodle#illustrator#artist#digital art#animal artist
1K notes
·
View notes
Text
If I knew how to code and model…
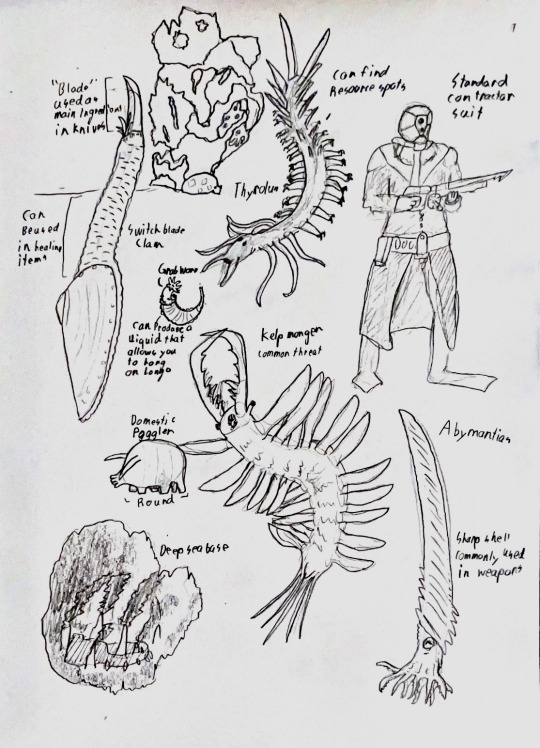

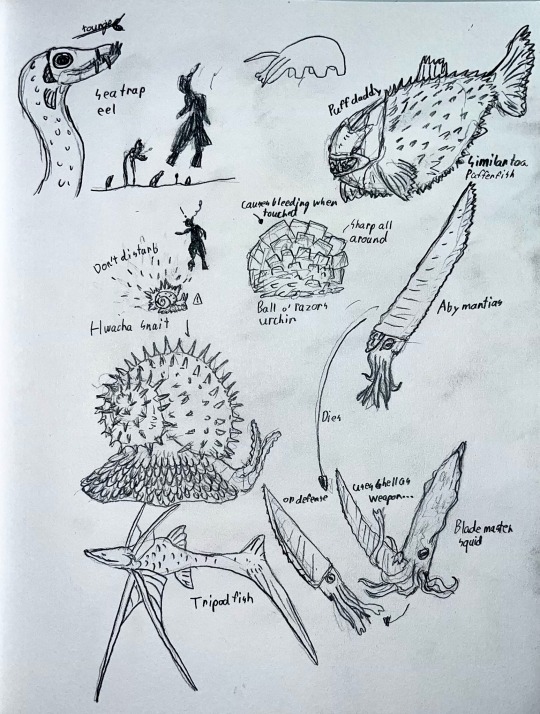
#speculative biology#speculative ecology#speculative evolution#speculative zoology#traditional sketch#traditional art#paleontology#paleontologist#spectember#paleo art#deep sea#video game concept#sketchbook#sketches#creature art#grappling#deep sea fauna#paleoblr
30 notes
·
View notes
Text





Have you ever heard of the 'Burgess Shale'? 😯 It's a goldmine of Cambrian fossils , a wonderland that is home to the most exceptional fossils from this era. Swipe through this post ➡️ to unearth the mysteries and ancient wonders that the Shale captured in its stones🪨 . To witness more of the prehistoric dance of animal life 🦕, join us on this monthly series 'Zoofabulous Time Trek'.
📸Image credits:
Image 1 - Olivia Little/ Freshdaily
Image 2- The Smithsonian National Museum of Natural History’s Burgess Shale Fossil Specimens
Image 3 - James St. John/ Wikimedia Commons
Image 4 - R. Gaines/ Semantic Scholar
Image 5 - The Burgess Shale Geoscience Foundation
#burgess shale#cambrian#cambrian explosion#fossils#paleo#paleontology#geology#zoology#animal#wildlife#fauna#prehistoric#education#science#science facts#study blog#discover#scicomm#research scientist#explore#earth
31 notes
·
View notes
Text


The Gorgonopsid Inostrancevia!
#art#my art#Permian#paleoart#paleo art#paleontology#paleontology art#paleontology sticker#Gorgonopsid#inostrancevia#Permian fauna#therapsid#paleo party
33 notes
·
View notes
Text
PALEO PIONEERS (VOL 1) IS OUT!



What could be better than dinosaur camp at the Natural History Museum? Being able to go back in time and experience prehistoric flora and fauna, of course!
From waxing philosophical over some of the earliest life in Earth's history all the way to playing tag in the age of the dinosaurs, Pioneers Miller, Aster, and Lucerne are going to maximize the amount of fun they have in the world of paleontology… OR FOSSILIZE TRYING!
VOLUME ONE of an all-ages comic series for both new discoverers of dinosaurs and long-term paleo enthusiasts alike!
GET IT HERE
#cecil draws#paleo pioneers#paleoart#paleo art#paleontology#comic zine#comic art#original comic#webcomic
270 notes
·
View notes
Text
⋯ paleontology themed id pack !!

names ⁘
fossil ⋄ trilo ⋄ dino ⋄ cambria ⋄ amber ⋄ nuna ⋄ shale ⋄ marrella ⋄ archo ⋄ azhdarcho ⋄ strata ⋄ phyla ⋄ devon(ia) ⋄ mosa ⋄ darwin ⋄ finch ⋄ kimberella ⋄ pteraspi ⋄ stega ⋄ raptor ⋄ burgess ⋄ holocene ⋄ lichida ⋄ luca ⋄ zircon ⋄ ediacara
trilo: as in trilobite | cambria: the cambrian period | amber: where many insects are preserved | nuna: one of the oldest supercontinents | archo: as in archosaur | azhdarcho: as in a group of large pterosaurs | strata: as in stratigraphy | devonia: the devonian period | finch: darwin's finches | stega: as in stegosaur | burgess: the burgess shale, british columbia | lichida: a group of trilobites | luca: last universal common ancestor | zircon: a crystal that can help search for early microbial life | ediacara: the ediacaran period

pronouns ⁘
paleo/paleo ⋄ fossil/fossils ⋄ dig/digs ⋄ dino/saur ⋄ dino/dinos ⋄ geo/geology ⋄ phy/phylum ⋄ tri/trilobite ⋄ bone/bones ⋄ mammoth/mammoths ⋄ flora/fauna ⋄ zo/zoa ⋄ tetra/tetrapod ⋄ evo/evolve
titles ⁘
The Most Ancient ⋄ (prn) Who Swims with Ichthyosaurs ⋄ The Fossilized One ⋄ (prn) Who is Encased in Stone ⋄ The Ancestor of Life ⋄ The Driver of Evolution
system names ⁘
the burgess shale ⋄ the fossil dig ⋄ the cycling lands ⋄ prehistoric pack ⋄ mesozoic menagerie ⋄ the index ⋄ heart of ediacara
As a certified paleontology nerd with a tattoo of a cheirurus trilobite and an anomalocaris.... this was absolutely necessary. -Iris (she/her)
credit: 1, 2
#cosmozoa— id pack 🌌#id pack#endo friendly#mogai friendly#name ideas#name suggestions#pronoun ideas#pronoun suggestions#system names#titles#npt#npts#npt list#npt ideas#npt suggestions#title suggestions#name list#neopronouns#noun names#unique names
80 notes
·
View notes
Text

DINOSAUR ID PACK
NAMES ⌇ amber. andrew. archie. archo. aroara. azhdarcho. bailey. barney. bowser. bronto. burgess. cambria. cera. charlie. cory. daemon. darwin. dee. devon. devonia. dina. dino. dinora. draco. eddie. ediacara. eli. felix. finch. finn. fleet. fossil. grover. holocene. huxley. jade. kaisen. kermit. kimberella. leo. lichida. luca. marrella. max. mite. mosa. nuna. oscar. phyla. pteraspi. quetzal. raptor. rex. rexie. reximus. rexley. rhino. roardin. roarke. rocky. ronaldino. roryn. roy. sage. sara. scout. shale. spike. stega. strata. tank. theodore. thesaurus. thumper. trexler. trilo. ty. verdi. ziggy. zircon.
PRONOUNS ⌇ bone/bone. cera/cera. cera/top. cerat/cerat. ceratops/ceratop. claw/claw. dig/dig. dino/dino. dino/saur. dinosaur/dinosaur. don/don. evo/evolve. flora/fauna. fluff/fluff. foss/fossier. fossi/fossi. fossil/fossil. geo/geology. hx/hxm. hy/hym. jur/jurassic. mammoth/mammoth. mim/mim. mimu/mimus. mimus/mimus. paleo/paleo. paleon/paleon. phy/phylum. plush/plushy. pteryx/pteryx. rapt/rapt. raptor/raptor. rawr/rawr. rex/rex. roar/roar. saur/saur. saur/saurus. saurus/saurus. scale/scale. shx/hxr. shy/hyr. spino/spino. teeth/teeth. tetra/tetrapod. thxy/thxm. thy/thym. tri/trilobite. tyra/tyra. tyrant/tyrant. tyras/tyrant. ven/ven. vena/vena. venator/venator. zo/zoa. ⭐ . 🌋 . 🍖 . 🦕 . 🦖 . 🦴 .
#⭐️lists#id pack#npt#name suggestions#name ideas#name list#pronoun suggestions#pronoun ideas#pronoun list#neopronouns#nounself#emojiself#dinokin#dino therian#dinosaurkin#dinosaur therian#paleocore
65 notes
·
View notes
Text








Last year, I created a trading card game concept featuring local paleo fauna of different formations. I want to produce a professional set this year and maybe an English edition as well.
The German trading cards concepts and pins will soon be available again in my online shop.
#paleoart#paleontology#scince illustration#digital illustration#trading cards#dinosaurs#basal tetrapods#synapsids#pterosaurs#natural history#archaeopteryx#dimetrodon
665 notes
·
View notes
Text
Palaelodus, the Swimming Flamingo
Another post for fossil flamingos since I just finished the page yesterday.
Palaelodus is a genus of early flamingo relative belonging to the family Palaelodidae, which can be aptly described as swimming or straight-billed flamingos.


Palaelodids are the sister family to Phoenicopterids, the modern flamingos, and already show a bunch of similarities to them like a deeper bill, long legs and neck and saltglands. Other elements meanwhile are more similar to grebes, which are the closest living relatives to flamingos. This has caused some debate on how Palaelodus would have behaved. Early research proposed that they may have been early filter feeders, but a closer look at the bill shows that they lacked some adaptations that hold the keratinous lamellae in flamingo beaks. Consequently, it has since then been suggested that they fed on insect larvae and snails that live in salt lakes, which may have been an early step for phoenicopteriforms to become filter feeders. The next question regards their legs. Early on people suggested they may have been divers, but that simply didn't make sense due to how strangely long they are. Others suggested wading habits, but their feet are strangely stiff. Mayr's hypothesis seems to make the most sense, arguing that they were swimmers, using stiff webbed feet as paddles.
Megapaloelodus goliath (=Palaelodus goliath?) by Scott Reid

Despite their very different lifestyle, Palaelodus actually had similar habitat preferences to flamingos, inhabiting salty and brackish lakes and having nearly global distribution. The most well known species, Palaelodus ambiguus, Palaelodus gracilipes and Palaelodus crassipes were incredibly common in Oligocene to Miocene Europe, especially France and Germany, the former country having over a thousand bones of them. In France, all three species (which some have suggested is only a single species of varying size) coexisted with the large Megapaloelodus goliath and the true flamingo Harrisonavis.
Images by Alphynix, Ceratops Yuta's Dinosaur Travel Blog and Torres et al. 2015



Outside of Europe, fossils from Brazil are known and tentatively assigned to Palaelodus ambiguus as well. Indetermined palaelodids are known from the early Oligocene of Egypt (Jebel Qatrani Fm), one species named Palaelodus kurochkini stems from Mongolia. Three species are known from Oceania. Palaelodus pledgei and Palaelodus wilsoni are the smallest and largest species respectively, having been found in the Oligocene to Miocene deposits of the Lake Eyre Basin in Australia. A single species, Palaelodus aotearoa, is known from the paleo-Lake Manuherikia in New Zealand, known for its extensive fauna of waterbirds. Much like the European species, the two Australian taxa coexisted with true flamingos, Phoenicopterus novaehollandiae and Phoeniconotius eyrensis.
Palaelodus cf. ambiguus by Felipe Alves Elias, Lake Eyre flamingos by Frank Knight and Palaelodus aetearoa by Tom Simpson



So all things considered, palaelodids and Palaelodus specifically did really well during the Oligocene and Miocene. What happened then is not known as they just disappear from the fossil record, unlike flamingos. Well, mostly disappear from the fossil record. There is one exception. A single isolated bone initially assigned to Palaelodus wilsoni was discovered at Cooper Creek in Australia, indicating an age of no more than 200.000 years. Since then its been suggested that it may not be P. wilsoni or even Palaelodus at all, but from the most recent publication I've seen on it the idea that its a palaelodid is still maintained. I am unsure on this one, as it is a massive gap in the fossil record, but there would be something tragic about it. Assuming that palaelodids survived in Australia, they would have been at home in a place that for a long time was a paradise for flamingos with salt lakes covering whats now the Tirari Desert. And then, during the Pleistocene when Australia got more arid and the lakes dried up, the former diverse flamingo fauna was gone for good, leaving Australia (and Antarctica) the only continents without flamingos, all because of some relatively recent climate change.

As always this is brought to you by me going down a wiki editing rabbit hole, the final result of which can be found here Palaelodus - Wikipedia. That article has all sources and information and then some more stuff that I didn't go in here (like the history and some anatomical lingo) Chances are that Megapaloelodus will be next, at which point I'll have written the pages for all fossil phoenicopterids and palaelodids.
#Palaelodus#Palaelodidae#Flamingos#Phoenicopteridae#fossils#flamingo#wikipedia#wikipedia editing#Palaelodus ambiguus#paleontology#palaeblr#long post
273 notes
·
View notes
Text
Commissions will stay their current price for 3 weeks!
I found the amount of effort and time I put into each commission felt far more than what I got out of it and it cannot sustain food spending.
Especially due to the low commission rates– I'd prefer to have low prices but many commissions so more people could get the chance to commission me but- unfortunately that is not doable.
I will ensure that I have lower cost commissions -- under $10 -- still, however this will exclusive be for lower effort/ single character sketches.
Thank you for understanding ! Please I still encourage you to commission me in this cheaper range while you can!
Treat it as a donation to me- the art in return is my gift back to you.
Examples of my art below:










I can draw people of all body types, ethnicities, etc --including scars--
creatures --spec evo, extant, and extinct..including paleo!--
I'm working on drawing more mechanical things such as robots but currently I specialize in fauna
I'd love to have more commissions of black and indigenous characters for example!!!
--and if you know this account at all you know this is a System/OSDD and DID safe space- yes, I can draw your alters--
#gordon post#commissions#I know these examples might be quite cookie cutter but I really would love to explore more characters and scenes given the chance!#Im not the fastest illustrator when it comes to commissions but this is often due to the fact I will study and ensure my ability to draw it#no matter what#I even did that with my commission for a utahraptor and I know quite a bit about dromaeosaurids#your satisfaction is quite important to me (directed to you the general client)
12 notes
·
View notes
Note
Oh that sounds fun. Personally an issue I always had with Pern's fauna was it felt like Anne never cared that much about making dragons feel like they belong in the same clade as Wherries and Tunnel Snakes outside of a passing reference. Like, why are Dragonets the only species to evolve the five color sex breakdown, or teleportation and telepathy? Imagine if there was a species of dragon-like fliers who could weakly go between (maybe they appear in the same place they left from as a way to dodge an attack), or if the five colors was ancestral to a major clade of "vertebrates," or a species of tunnel snake that hunts with a telepathic attack to stun its prey.
Oh she totally didn't care and considering Anne is also the same person who had cheetahs as man-eating dragon-hunting apex predators I think we really can't trust that she knew anything about animals either way lol
As for all those things you mentioned, I hc (and am gonna show in my "wildlife of Pern" art) that the five-color sex morphs are the default among Pern's vertebrates, it's just that the sexual dimorphism (penta...morphism...?) is more noticeable in dragonkin, and as for the telepathic and between-going animals, welp. they got Threaded 😔 (and I'm still gonna design them because fuck it! Pern paleo art!)
#dragonriders of pern#I like the ''go between and reappear in the same place''!. I think it would still make sense that the species would go extinct#to explain their absence. because with predators yeah you're gonna dodge the attack but with Thread you're still#returning from between and landing onto infected ground
14 notes
·
View notes
Text
A new pterosaur specimen from the Lower Cretaceous Yixian Formation of Liaoning Province, China: The oldest fossil record of Nurhachius
Published 28th June 2023
A new fossil specimen of the Istiodactylidae Nurhachius from the Liaoning Province.
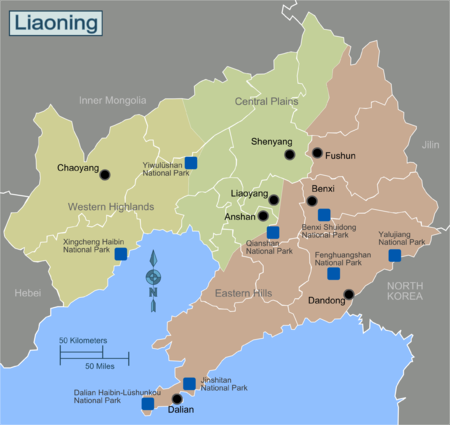
Image of the Liaoning Province, from Wikitravel.

Nurhachius ignaciobritoi described 6th October 2005.

Nurhachius luei holotype (BPMC-0204), described 19th September 2019.
Nurhachius is an almost mature pterosaur with an estimated wingspan of 1.6 m and is geologically the oldest known occurrence of Nurhachius. The istiodactylidae is a new component of the pterosaur assemblage of the Jingangshan Member, and provides new information on the pelvis and hindlimb of istiodactylids and the palaeobiology of this clade. A great discovery that provides more information for the pterosaurs in China.
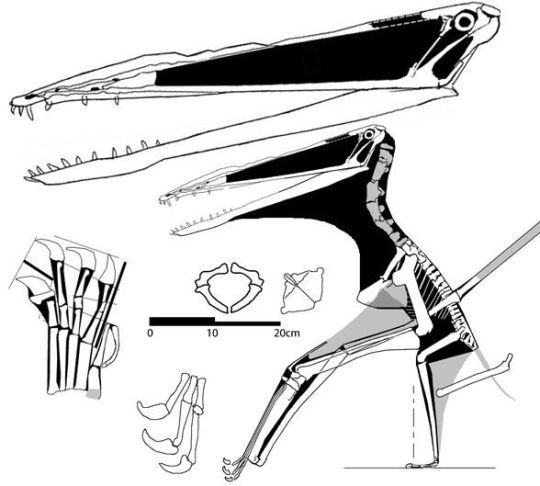
Skeletal reconstruction of Nurhachius, from:
note that this reconstruction may not be reliable or accurate and may be a speculative/innovative creative vision of Nurhachius' appearance (note added in response to feedback, thank you! I'll make sure to include disclaimers/warnings in the future for less trustworthy sources)

Paleo art of Nurhachius, on Wikipedia.
Source:
read the paper for more information.
36 notes
·
View notes
Text
South West really is fucking best
Just within 30 mile radius of where I live we had Natives, Mormons, Cowboys, Miners, Confederates, Mountain Meadows Massacre, Spanish Trail, Mormons vs Mexicans, Silver boom towns that saw legends such as the prosecuter in the OK Corral shooting case and one of the men responsible for turning Las Vegas from a mining camp to a boom town, we got an unknown number of mines, we got peaches and pecans and apricots and decent agriculture, we got all manner of flira abd fauna that'll screw up your day, we got dinosaur bones and dinosaur tracks, we got mastadons and obsidian, we got paleo rock art nearly everywhere the natives lived, we got endless trails of sand and rock, we got cliffs with fossil beds going back 400 million years, we got an entire mountain comprised of red sand and red sandstone, we have several monumental canyons, we had thousands of tons of radioactive fallout, we have rocket sled test facility where they used to test ejection seats, two airports and so much more.
I don't want to live elsewhere.
14 notes
·
View notes
Text









Imagine 💭a world where evolution tripped on a power line.💡 That's the Cambrian! Before dinosaurs, 🦖 before fish, 🐟 even before shells,🐚 the Cambrian seas flourished with life unlike anything we know. To dive 🥽deeper into its fascinations, hop onto this 2nd trip of the 'Zoofabulous Time Trek' with us where we'll be explorin' 🔎 the Cambrian period!
#zoology#animals#cambrian#cambrian explosion#paleo#paleontology#education#science#science facts#discover#scicomm#paleozoic#wildlife#fauna#animal#hallucigenia#anomalocaris#opabinia#wiwaxia#pikaia#trilobite#geology#archaeology#fossils#evolution#prehistory#biology#pokemon#extinct animals#extinction
23 notes
·
View notes
Text
Paleontological Biogeography of Niur
The unique characteristics of Niuri fauna (and to a lesser extent flora) have been shaped by the geology and isolation of the islands, as they moved through time and space, as well as the unique conditions surrounding them.
Already at the start of the Paleocene, the landmass that would one day form the Niuri islands was already somewhat loosely attached to the South American mainland. Although it fluctuated with sea levels, for most of this period only an isthmus connected the two, and this prevented total faunal exchange from occurring, allowing, even then, isolation enough for the Niuri fauna to diversify separately from their South American relatives.

The Niuri landmass was climatically quite different from the neighboring region of South America, which additionally was a factor limiting the free transfer of organisms back and forth between the landmasses. Even so, outside of insects and some other groups of invertebrates, the fauna of the Niuri landmass, and for the most part the flora, was largely indistinct from that of South America until the middle of the Paleocene, differing mainly in the varying amounts of taxa found rather than the types. Towards the middle Paleocene , however , a low lying area in the middle of the Niuri landmass began to sink and was flooded, first as a large lake and then shortly after as a bay.
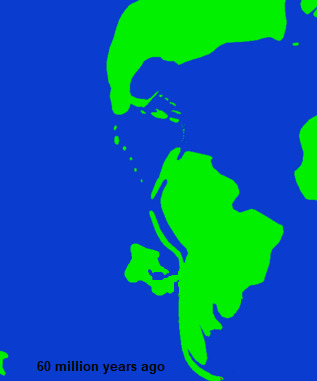
In tens of millions of years this bay would become part of the Faln sea, but even from its formation it began to change the life in the surrounding area. The most immediate effect was the change in marine fossils, as the shape of the land around the bay isolated it from the greater Pacific and also provided a unique habitat within it. Additionally, it triggered a further differentiation of the habitat of the Niuri landmass from the South American mainland, especially in the region of the isthmus. Finally, it is believed to have contributed to a broad shift in the climatic stability of the landmass as a whole, as prior to this point, the portion of Gondwana that would one day split of that would eventually become the Niuri landmass was subject to a high degree of climate instability , particularly in inland environments, and tended to support a very low diversity of megafaunal animals.
Beginning with the formation of the Paleo-Faln Bay the middle Paleocene the Niuri climate would change to instead become remarkably stable and consistent during the Neogene, much more so than neighboring continents or indeed most of the world. That said any semblance of stability during the last half of the Paleocene was rather abruptly interrupted by the end of that period.
The Paleocene-Eocene Thermal Maximum appears to have abruptly altered the environment of the Niuri landmass, and it triggered a rapid shift in its faunal makeup. Taxa which had previously been common throughout both Niur and the adjoining regions of South America disappeared or were heavily reduced in Niur, while some groups that all but disappeared in mainland South America continued to flourish in Niur. In the Ypresian, the first period of the Eocene, for the uniquely Niuri fossils of mammals, reptiles, and birds begin to make up a substantial degree of the Paleo record, by the beginning of the Middle Eocene the fossil taxa of Niur is almost wholly distinct from South American forms. It's also around this time when the last connection to mainland South America disappeared, and from then on Niur was an island landmass, drifting to the northwest on its Continental plate.
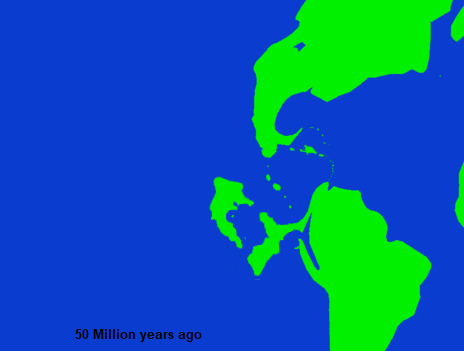
While it last was directly connected to South America no later than 40 million years ago, Niur was still close enough that birds, plants , some insects and bats could readily cross the gap. Even so this isolation was complete enough that from then on all future introductions of new fauna to the Niuri Islands are termed as distinct events. Even prior to becoming truly isolated, the faunal interchanges of Niur and South America in the Paleocene and Early Eocene are also considered to be distinct events, namely the First and Second South American introductions. The First Introduction refers to a period of time in the late Paleocene, where after some degree of distinction had already emerged in the islands some new faunal groups migrated over from South America. The more impactful Second Introduction occured immediately prior to the final severance, somewhere around 45-40 million years ago, perhaps caused by a temporary widening of the connection between the landmasses before it finally was buried under the sea. These new groups were subject to almost immediate, intense evolutionary pressures upon Niur's isolation, and those that survived rapidly diversified into new forms.
In the early Eocene, the western half of the Niuri landmass had been rendered an island and with the separation from South America, and these would retain separate faunal assemblages until the Oligocene, when changes in the landscape and sea level led to an interchange of animal taxa across the islands. The landmasses that composed these islands were heavily altered in position and shape, and are not to be confused with the two later proto-islands that would appear (along with Senika) later in the Neogene. The largest differences between these two assemblages were mammalian taxa, with birds and reptiles appearing to be less affected by the gap between them, although it's believed that the poisonous bird groups originated in the western island before also expanding into the eastern island as well. While both western and eastern islands included members of the initial mammalian assemblages, as well as those that arrived during the First South American introduction, the mammals that arrived in the Second and later Third South American (which occurred in the latest eocene or earliest oligocene, as a chain of islands allowed some taxa to island hop from northern South America) introductions were limited to the eastern island until the late Oligocene. Most prominent among the eastern limited animals were the Notoungulates, the Pyrotheres, and later the Astrapotheres and ameridelphian marsupials all of which would lead to unique Niuri taxa that are still extant. In addition to this, xenarthrans were present in the eastern island as well, although these remained species poor and went extinct by the end of the Oligocene. There are at least some teeth fossils that suggest that Litopterns may have briefly been present in the eastern island, in the mid Eocene.
Meanwhile the only major group of placental mammal to survive in the western island during this time period were the xenungulates (of the first South American introduction), which although initially being somewhat diverse and common in the early Eocene, afterwards appeared to have been in a long decline before disappearing in the Oligocene. The other mammals present in the western island were the various non-placental mammals, which along with the Niuri ameridelphians (Kaulodelphidae) , are collectively called "Koutou" in modern Niur, despite not being particularly closely related. These include the Gondwanatheres, and Polydolopimorphia, both present from the earliest Paleocene, and the Sparrassodonts, and the Xenodelphians from the First South America introduction. The Sparrassodonts were additionally present in the fauna that arrived during the Second South American introduction, and the new arrivals appear to have replaced the Sparrassodonts still present on the Eastern Island, although after the faunal interchanges in the Oligocene both Western and Eastern Sparrassodonts continued to flourish. Additionally, the Polydolopimorphia maintained high diversity on both western and eastern islands until the Oligocene, and all three modern families appeared nearly simultaneously on both islands before the faunal interchanges that unified the assemblages (although one extinct, enigmatic family that appeared to be endemic to the eastern island went extinct not long after).

Shortly prior to the Oligocene Faunal Interchange, there was one additional "introduction event" wherein, most likely via a single rafting event, salamanders were introduced to what is now Senika (which was the earliest modern island to differentiate itself clearly in the fossil record). In the Niuri paleontology this is called either the First North American Introduction or the Zero Suri Introduction, as its debated whether they rafted from North America or from the Suri islands. They would gradually spread during the rest of the Oligocene, before more heavily diversifying in the Miocene.
In the mid to late Miocene, the Suri islands, themselves subject to a long and generally less understood geological history, approached close enough to the east of Niuri that fauna and flora could exchange between these two island chains. The fossil record of the Suri islands is confusing, and this is further complicated by the fact that they have been subject to gradually increasing subsidence for the past 10 million years, which along with the rise of sea levels during the Holocene , has buried much of the former Suri landmass under the sea. This process of sinking was somewhat abated by a period of volcanism which created new land inbetween Suri and Niur, as well as contributed to the geological features of eastern Niur, that said much of this volcanism had dissipated by the Pleistocene.
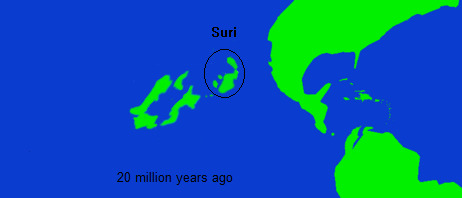
The first evidence of an interchange between Suri and Niur, actually appears in Suri, around 15-18 million years , when rather abruptly various distinctly Niuri plant pollen taxa appear almost simultaneously throughout fossil sites in Suri. This is followed shortly after by Niuri avian fauna, which although less attested appears to rather rapidly replace the indigenous Suri birds (although how complete this replacement was is hard to say, given the fragmentary record). The introduction of Niuri life in Suri seemed to trigger a wave of extinctions as well as spur the new evolution of Suri life.
Generally it seems that during the Miocene, most introductions between Niur and Suri tended to follow the pattern of Niuri life establishing itself in Suri, with little managing to travel the other way. Following the avifauna, Niuri Astrapotheres and Xenodelphians arrived in Suri, becoming firmly established during the late middle Miocene. Around this time, the two island chains reached their closest point of contact, after which Niur would continue to drift eastward at a greater speed than Suri. Even so, because of the lowered sea levels during the Pliocene and Pleistocene, and continuing volcanicism creating temporary, connective archipelagos, the faunal interchanges would continue, although in separate pulses. The last phase of the the first Suri introduction was the point at which Suri fauna, perhaps finally adapted to the pressures of the Niuri arrivals, managed for the first time to make the journey to Niur. Between 10 and 7 million years ago, a variety of Suri taxa became established in Niur. The first, and most widespread of these groups, were the two Suri families of salamander, which now accompanied the earlier cryptobranchid salamanders that had been introduced in the Oligocene. Additionally, for the first time, rodents found their way to Niur, the small mouse like members of the family Sminthidae (one of the only rodent families to survive in Suri) adapted to alpine habitats in Niur, and in present at least five species are known in the genus Subtilicus. The one megafaunal introduction, though, was the most impactful, the carnivoran feneraks. These catlike predators are part of the larger clade Felarimorpha which is either a suborder of Carnivora itself or an infraorder of Feliformia (although some phylogenetic analyses have proposed that Felarimorpha might instead be the sister taxa of all other carnivorans) . Feneraks make up one of the two living clades of Felarimorphs, although the first feneraks in Niur were part of a now extinct group, often called the Crested Fenerak, after the unusual protrusions that later members developed on their skulls. Crested fenerak gradually spread throughout much of Niur, and took the place of the sparrasodonts as top mammalian predators (and indeed, in many habitats, as top terrestrial predators in general). Crested fenerak would continue to evolve in Niur, while new groups, more closely related to modern fenerak, evolved in Suri. Niur and Suri drifted away from each other, the first period of interchange ended, with the fauna of both islands profoundly shifted.

The two island chains, for much of the Pliocene, existed largely in separation from each other, although birds and plants still managed to easily cross from one to the other. Niur, especially the southern parts of it, has many plant genera in common with Central America and Northern South America, and while its unknown exactly when they made the journey there, many groups present are believed to be relatively recent evolutionarily speaking, and therefore may have been the result of birds ferrying their seeds from the continents , to the southern parts of Suri, and then to Niur. Most prominently, the wide variety of cactuses in Niur, many of which are relatively closely related to species found in North and South America, is believed to be caused by such patterns. This might also be where many of the Solanaceae present in Niur originate from, and unless the result of an ancient introduction by people, almost certainly how the genus Capsicum arrived.
Both Niur and Suri experienced environmental and ecological changes during the Pliocene, and in Niur at least, this lead to the decline of the crested-fenerak in most locations other than what is modern day Nirou, by the end of the Pliocene. Rather than disappear all at once, they seemed to gradually decline area by area, at first in the south of greater Kaita, with their disappearance spreading from there until they were restricted to the northern plains of Nirou. This disappearance was historically tied to the onset of the Second Suri introduction, which is considered to have started shortly before the onset of the Pleistocene, as lower sea levels and renewed volcanic activity again created a bridge between Suri and Niur, and allowed the dispersal of the modern feneraks into Niur. However, more recent research suggests that the crested-feneraks began their decline before modern feneraks arrived, and in fact it was only in areas where they had already disappeared that modern feneraks were able to establish themselves, with the exception of northern Nirou, where crested-feneraks survived for much longer and coexisted with their modern cousins until eventually going extinct during the late Pleistocene. There are five species of fenerak alive today in Niur, and they probably evolved from at least somewhat semi-aquatic ancestor (as the water-fenerak is today still) which managed to island hop to eastern Niur, although its unclear if this was the result of one or two introductions. The record throughout much of the Pleistocene is ambiguous, and the exact history of the fenerak might be better elucidated by further genetic studies. The other group of Suri animals that is typically considered to be part of the second introduction were the peccaries, which live today in various forests on the eastern Islands of Niur. Although their choice of habitats is different (both in Niur and Suri they exclusively live in relatively cool forests) they seem closely related to the American peccaries, to the point where its a bit of mystery as to how they arrived in Suri in the first place. Compared to the fenerak which are usually apex predators or at least dominant in their environments, Niuri peccaries, while locally common, tend to be more marginal parts of their ecosystem, and even still mostly live in areas where they have less competition from other mid to large size herbivorous animals.
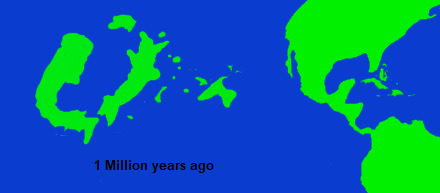
There is no exact boundary between the Second and Third Suri introductions, because low sea levels throughout the Pleistocene meant that there was an almost continuous potential for island hopping between Niur and Suri. Rather, the Third introduction is defined by its most famous member, the people of Niur, the felar-mesai, who first arrived in Niur from Suri, sometime in middle Pleistocene. Because of their beliefs regarding remains of the dead and genetics, most studies on Niuri paleoarcheology have focused on material culture, and because of this many dates regarding the earliest records of people in Niur are subject to massive ranges. Its also unclear if the earliest people in Niur were members of the same species as present day mesai, and the fragmentary record of remains is complicated by the fact that there is a high degree of genetic diversity within the present day mesai (much more so than modern humans), meaning that any species level determinations based on bone fragments are highly contested. Different methods of dating have suggested various results of when the first people in Niur arrived (or at least, the first member of the clade of felarimorpha to which modern mesai belong to), anywhere from 150,000 to 800,000 years ago, a time range of over half a million years. These inconsistencies might be explained by the "lost seeds" hypotheses, which views these early records as evidence not of permanent populations but rather temporary, small groups of mesai, which periodically would arrive in Niur during certain climatic periods, which corresponded to a spread of specific Suri plant taxa in Niur, and then disappear when the climate shifted again and those plants became less common, without ever becoming fully established in Niur. Even those that oppose this theory, tend to view the populations of Niur during this time as being sparse and likely subsumed by later arrivals. The period around 120,000-70,000 years ago is seen as the first clear period of continuous inhabitation of Niur, and appears to correspond with a new group of people arriving from Suri, or at least a new material culture spreading from there. Even so, population density was incredibly low, and subsequent migration from Suri in various pulses continued until around 20,000 during the last glacial maximum. The mesai population in Suri would decline considerably after this point, with the last evidence of living mesai dating to perhaps around 10,000 years ago, although there may have been a very small remnant population until as late as 8,200 years ago.

Along with mesai, at least in the later migrations, included various semi-domesticated Suri plants, which form the bulk of the other arrivals during the Third Suri introduction. By the time of the last glacial maximum, the volcanicism that had created island chains between Suri and Niur had mostly subsided, and the islands that were left gradually sunk into the rising sea levels of the Holocene. The last remnants of that volcanism are the hot springs and mountains of Senika, Nacre and the islands to its north. Despite the massive changes in climate and land, as well as the arrival of people, most of Niur's native fauna remains highly successful, and incredibly distinct. Groups that have long gone extinct elsewhere thrive there, and its unique ecosystems have so far avoided the fate of other isolated environments that has so often come about as the result of new introductions. Outside of some plants and a few insects associated with introduced agriculture, almost no invasive species have managed to take hold in Niur, and few if any native taxa have gone extinct in the last 10,000 years.
On the contrary, the robustness of Niuri ecosystems have leant some Niuri taxa the ignominious status of being invaders themselves, mostly a handful of insects and plants, aside from one specific incidence where one of the large Niuri ratites (the Upland Takako) become established in parts of Spain after a failed attempt to farm them. These large birds have proved to be a nuisance in Spain, as well as being environmentally damaging, as they will kill foxes or other mammals they view as a threat to their eggs or young, as well as domestic goats and sheep they view as competition for food. Attempts to control them via hunting have been partially successful, although the surviving population (mostly in the Pyrenees) is much more wary of humans and traps and is also notoriously dangerous, sometimes injuring or even killing hunters (in addition to sheep and goats).
3 notes
·
View notes
Text
COMING SEPTEMBER 1: PALEO PIONEERS VOL. 1!




What could be better than dinosaur camp at the Natural History Museum? Being able to go back in time and experience prehistoric flora and fauna, of course!
From waxing philosophical over some of the earliest life in Earth's history all the way to playing tag in the age of the dinosaurs, Pioneers Miller, Aster, and Lucerne are going to maximize the amount of fun they have in the world of paleontology… OR FOSSILIZE TRYING!
VOLUME ONE of an all-ages comic series for both new discoverers of dinosaurs and long-term paleo enthusiasts alike!
#cecil draws#paleo pioneers#paleoart#paleo art#original comic#comic zine#zine#stegosaurus#hallucigenia#paleontology
124 notes
·
View notes
Last Seen Blogs

380965837361
BananaDay
380965837361
BananaDay

blood-mocha-latte
and that's all eternity is

bledloyalty
I ain't ALWAYS
380965837361
BananaDay