#Testate amoeba
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
Total funding amounts to $125.3M.
Text
youtube
Look at this testate amoeba (that is, an amoeba that has a little shell around it), just crawling along by sticking its pseudopods (tentacles) out of holes in the test/shell. The light gleams off its shell, making it look so beautifully ethereal and alien...
29 notes
·
View notes
Link
#Pobitora wildlife sanctuary#Testate amoeba#Grassland#Keystone species#Conservation#Greater one horned rhinoceros
1 note
·
View note
Text

A few days ago, scientists published an exciting discovery in the icy waters of the Arctic Ocean: Katarium polorum, a new cold-loving amoeba that’s turning heads in the world of microbiology. Discovered by Marcel Dominik Solbach, this tiny organism is only about 15 micrometers in diameter, but it’s packed with some pretty impressive survival strategies for living in one of the most extreme environments on Earth.
What makes Katarium polorum stand out? For starters, the organism is encased in an organic shell, with a small round opening. From this opening, a threadlike extension of cytoplasm often pushes out, forming delicate, branching pseudopodia. These slender extensions are used to explore the surrounding environment—almost like tiny feelers on a quest for food or new territory. Interestingly, these amoebae don’t seem to be ones for settling down. Instead of attaching to surfaces like many other microorganisms, they float freely in their surroundings, continuously extending and retracting their pseudopodia, as if navigating an ever-changing landscape.
The primary diet of Katarium polorum seems to be diatoms, though it has also been seen feasting on bacteria. But it’s not just the way it moves or feeds that’s captivating scientists—it’s also how it behaves as it ages. In older cultures, the amoeba begins to clump together, sometimes forming clusters of several or even dozens of cells. Among these groupings, researchers have found enormous, multinucleate cells—known as "monstrosities"—offering a fascinating look at the amoeba’s biological complexity. These giant cells seem to represent a key piece of the puzzle when it comes to understanding how this organism thrives in the frigid waters of the Arctic and Antarctic Oceans.
Another intriguing feature of Katarium polorum becomes visible in these older cultures: a distinct layer of reflective granules or crystals that form a horizontal band within the cell’s cytoplasm. Despite these changes, the amoeba remains active, continuing to feed, move, and divide. It’s as if it is constantly adapting to its harsh surroundings, constantly evolving to survive the coldest places on the planet.
The discovery of Katarium polorum opens new doors in our understanding of life at extreme temperatures, shedding light on how microscopic organisms can not only survive but thrive in conditions that would be unthinkable for most life forms.
For the curious and the scientifically minded, you can read more in the full research paper here: https://doi.org/10.1111/jeu.13071
#nature#biology#microbiology#microorganisms#protists#testate amoebae#zoology#amoeba#life science#science#protistology#microscopic organisms#microscopic#microscope
2 notes
·
View notes
Text
Peat bogs, which cover 3% of the world’s land surface, are special places. While historically often considered as worthless morasses, today they are recognised as beautiful habitats providing environmental benefits from biodiversity to climate regulation. However, they are threatened by drainage, land reclamation for agriculture and peat cutting for fuel, which has significantly reduced the extent and condition of these ecosystems on a global scale. Bogs are fragile and sensitive to change, whether by human hands or by processes such as climate change.
A less well known aspect of bogs is their remarkable archaeological potential. In their undisturbed state at least, bogs are anoxic (oxygen-free) environments due to their saturation. These conditions are hostile to the microbes and fungi that would normally decay organic material such as the remains of plants, which are the principal constituents of the peat. The same anoxic conditions also offer protection from decay for organic archaeological remains. The vast majority of objects and structures used by our ancestors were made from organic materials (in particular wood). These are normally lost on dryland archaeological sites but can be preserved in peatlands.
But archaeology is only part of the story these environments have to tell. They are important archives of the past in other ways: the layers of moss and other vegetation that make up peat are themselves immensely valuable as archives of past environments (palaeoenvironments). The manner in which peat accumulates means that the deposits have stratigraphic integrity, meaning that contained within each layer can be found macroscopic and microscopic remains of plants and other organisms that shed light on landscape change and biodiversity on timescales ranging from centuries to millennia. The high organic content of peat means that these records can be dated using the radiocarbon method.
The best known such records are probably pollen grains which provide evidence of past vegetation change. But evidence from other organic material can be used to reconstruct other past environmental processes. For example, single-celled organisms called testate amoebae, preserved in sub-fossil form, are highly sensitive to peatland hydrology and have been extensively used in recent years to reconstruct a history of climatic changes. Meanwhile, fossil beetles can tell us how the biodiversity and nutrient status of a peatland has altered over time.
The potential of bogs to preserve both environmental and archaeological records means that they can be regarded as archives of “hidden landscapes”. The accumulating peat literally seals and protects evidence of human activity ranging from the macroscopic (in the form of archaeological sites, artefacts and larger plant and animal remains) through to the microscopic (pollen, testate amoebae and other remains) material that provides contextual evidence of environmental processes.
19 notes
·
View notes
Text
" International Symposium on Testate Amoebae(ISTA, ブロック大学, カナダ)2025年6月23日〜27日)“にて口頭発表を行います。 "Food-Dependent Behavioral Changes and Pseudopodial Dynamics in Lesquereusia spiralis”
Norihiko Kamaya¹, Toshiyuki Nakagaki², Atsushi Taniguchi², Yukinori Nishigami² ⑴ Graduate School of Life Science, Hokkaido University, Sapporo, Hokkaido, Japan, ⑵ Research Institute for Electronic Science, Hokkaido University, Sapporo, Hokkaido, Japan
------- The behavior of testate amoebae plays a significant role in nutrient cycling within their environment. However, their behavioral characteristics, especially those related to environmental cues and foraging strategies, are still poorly understood. Lesquereusia spiralis is a freshwater testate amoeba commonly found in nutrient-rich environments, where it feeds on filamentous algae such as Spirogyra. Previous studies have suggested that it may also consume small metazoans like nematodes, indicating a broader and more varied range of foraging strategies. In this study, we investigated the movement and pseudopodial activity of L. spiralis under two different experimental conditions. One condition featured a conditioned URO medium where Spirogyra had grown, while the other condition used a fresh URO medium. We observed the behavior of the L. spiralis in each medium. In the conditioned medium, L. spiralis frequently extended multiple pseudopodia in various directions and often remained stationary while actively exploring the area around the cell. These pseudopodia were short-lived and dynamically adjusted. In contrast, in the fresh medium, the amoebae extended fewer pseudopodia, maintained them for longer periods, and moved more steadily along linear paths. Overall, locomotion was faster and covered greater distances in the fresh medium. These results suggest that food-derived substances influence the movement patterns and the number of pseudopodia in L. spiralis, potentially promoting more efficient foraging in this species.
2 notes
·
View notes
Text
Test
In biology, a test is the hard shell of some spherical marine animals and protists, notably sea urchins and microorganisms such as testate foraminiferans, radiolarians, and testate amoebae. The term is also applied to the covering of scale insects. The related Latin term testa is used for the hard seed coat of plant seeds.

2 notes
·
View notes
Text
Seeds through time
By Addie and Mina, PLE Wetlanders Despite extreme physical exertion in the bog yesterday, Bob jumped out of his night-time waders to wake us up with highly animated morning lectures on peatland paleoenvironmental archives and wetland change over time. Excitingly, he discussed his own research examining how vast datasets on modern communities of testate amoebae (protists) in peatlands can be used…

View On WordPress
2 notes
·
View notes
Text
Can't wait for you to get to Testate Amoebas!!
Guys I can't draw another fucking anthro mushroom without chewing my hands off, I am switching out fungus for bacteria this year's Funguary - I think I'll call it Microbuary (working title)
6K notes
·
View notes
Text
Wk 16, 9th of May, 2024 Research
Findspot
findspot: the place where an archaeological object has been found or a place where an object is found.
From the text: A multi-proxy palaeoenvironmental investigation of the findspot of an Iron Age bog body from Oldcroghan, Co. Offaly, Ireland by Gill Plunkett...
In 2003, the remains of an Early Iron Age bog body, known as ‘Oldcroghan Man’, were recovered during the cutting of a drainage ditch in a bog in the Irish Midlands. Only some fingernails and a withe fragment remained undisturbed in situ in the drain face, providing the sole evidence for the original position of the body. A detailed reconstruction of the depositional context of the body has been undertaken through multi-proxy analyses of a peat monolith collected at the findspot. The palynological record shows that the surrounding area was the focus of intensive human activity during the Later Bronze Age, but was largely abandoned during the Bronze Age–Iron transition in the mid-first millennium BC. In the mid-4th century BC, a bog pool developed at the site, evidenced in the stratigraphic, plant macrofossil, testate amoebae and coleopteran records. Plant macrofossil and pollen analysis of peat samples associated with the fingernails suggests that the body was deposited in this pool most likely during the 3rd century BC. The absence of carrion beetle fauna points to complete submergence of the body within the pool. Deposition occurred shortly before or around the time that the surrounding area again became the focus of woodland clearance, as seen in the extended pollen record from the peat monolith. This period corresponds to the Early Iron Age in Ireland, during which renewed cultural connections with Britain and continental Europe can be seen in the archaeological record and widespread forest clearance is recorded in pollen records from across Ireland. The palaeoenvironmental results indicate, therefore, that the demise of Oldcroghan Man took place at a pivotal time of socio-economic and perhaps political change.
This paper describes the results of a multi-proxy palaeoenvironmental study conducted on a peat monolith collected from a section adjacent to the findspot of Oldcroghan Man. Using pollen, plant macrofossil, testate amoebae and coleopteran analyses, we reconstruct the landscape history of the surrounding area in the centuries bracketing the burial, as well as detailed local environmental conditions at the time the body was deposited in the bog.
From the text: THE FINDSPOTS OF BRONZE AGE METALWORK IN LOWLAND ENGLAND: A NEW FIELD STUDY by David Dunkin, David Yates and Richard Bradley...
This paper is concerned with the deposition of Middle and Late Bronze Age metalwork in three study areas extending from the coast of south-east England to the edge of the Fenland. It considers where hoards and single finds have been discovered and is based on inspection of over 300 findspots on the ground. It investigates regions in which tools and ornaments are common, but also extends to those with a stronger representation of weapons. While rapiers, swords and spears are found in rivers, terrestrial finds often come from sites close to springs, aquifers and confluences and favour areas with fresh water rather than salt water. Others are from more prominent locations on high ground or in low-lying positions like dry valleys. We suggest that such places played a role in a tiered cosmology. Metalwork was deposited where the underworld or the sky came into contact with the landscapes inhabited in daily life.
From the text: Staying local – experiencing local landscapes and the potential of hidden stories by Steven Timoney...
This article considers some of the opportunities increased visitation to recreational areas presents in terms of engaging residents and visitors with local heritage in the form of Historic Environment Records (HERs). There are countless potential ‘hidden stories’ related to cultural heritage sites and artefacts within palimpsest landscapes. These sites and artefacts present opportunities for alternative narratives of place to be negotiated, creating links to the past in the present, and between the exotic and the familiar. The changing approach to landscape practices resulting from COVID-19 restrictions presents new opportunities to engage people with these hidden stories, linking places near with far-off lands across time and space. It also encourages new connections with place that have the potential to (co-)create new narratives; increasing engagement, as well as correlating with pressing issues centred around the climate crisis and the urgent need to reduce travel to achieve Net Zero targets.
Taking Kinnoull Hill Woodland Park on the outskirts of the Scottish city of Perth as its focus, this article considers some of the opportunities this increased visitation presents in terms of engaging residents and visitors with local heritage. Within the landscape are countless potential ‘hidden stories’ related to cultural heritage sites and artefacts as part of palimpsest landscapes. These sites and artefacts present opportunities for alternative narratives of place to be negotiated, creating links to the past in the present, and between the exotic and the familiar. Heritage is often something that people travel to experience, and previously what was local may have been undesired or unknown (Lew Citation2018). Broadly speaking, outdoor recreation spaces saw an increase in activity resulting from varied COVID-19 lockdown measures (Bustad, Clevenger, and Rick Citation2023, 90). The restrictions on movement also provided the opportunity for explorations of ‘urban green spaces and the (re)formation of perspectives and practices attached to these’ (King and Dickinson Citation2023, 112). The changing approach to landscape practices resulting from COVID-19 restrictions presents new opportunities to engage people with these hidden stories, linking places near with far-off lands across time and space. It also encourages new connections with place that have the potential to (co-)create new narratives, increasing engagement; as well as correlating with pressing issues centred around the climate crisis and the urgent need to reduce travel to achieve Net Zero targets.
Chapter: Perceptions of landscape
The concept of landscape has evolved from its origins as landschap, used to refer to an area of land (Antrop Citation2013, 12), to what Wattchow describes as ‘a classic trans-disciplinary concept’ (Citation2013, 87). It is a complex construct that need to be interpreted, a reflection that social realities are constructed rather than innate (Atha et al. Citation2013, 2). As such, landscape does not just exist. Instead, landscape has been variously conceived as both a spatial and cultural entity (Brace and Geoghegan Citation2010, 287). They are texts to be read (Duncan and Duncan Citation1988); something that favours the visual, though the conscious and subconscious actions of individuals identify what is seen and how it is understood (Antrop and Van Eetvelde Citation2017, 62). It has been considered a dualism between its material reality and ideal representation (Timms Citation2008); landscape as a verb as well as a noun (Leyshon and Geoghegan Citation2012, 240). Though the separation of the cultural and the natural is not so clear cut:
As the familiar domain of our dwelling, [landscape] is with us, not against us, but it is no less real for that. And through living in it, the landscape becomes a part of us, just as we are a part of it (Ingold Citation 2000, 191).
Bound up within discussions of landscape are ideas around place, identity, and attachment. Aspects of identity then can have a geographical component. Place-identity refers to the way people use place as an influencing factor in the construction of identities (Proshansky, Fabian, and Kaminoff Citation1983). Tuan (Citation1974) saw space given meaning through the development of identities connected to landscape – what he defined as ‘topophilia’ (a love of place), with stronger attachments to place developing over time. Massey uses the term ‘a progressive sense of place’ (Massey Citation1994) to reflect the idea that the character of places is constructed by the flows of people and ‘things’ in and out of a space. Place attachment (Altman and Low Citation1992; Devine-Wright Citation2009) creates a sense of belonging with a spatial context in the sense of being in place.
Chapter: Perceptions of heritage
The purpose or role of heritage has been widely debated. Perceived distinctions between tangible remains and ‘objective’ history, have been compared to subjective, populist accounts which have been defined as heritage (Lowenthal Citation1998). The process of heritage is a selection of elements in the present that can be attributed to or provide connection with the past (Smith Citation2006; Tunbridge and Ashworth Citation1996). It has been argued, then, that these processes have facilitated a continued separation between archaeology and heritage in terms of management and practice, enabling archaeologists to be ‘very successful in protecting what they perceive to be their database’ (Smith and Waterton Citation2009, 1).

Bronze Age Gold from the Heyope hoard, The Museum of Wales online database in the archive Ancient Wales

Bronze Age Gold and tarnished metal from a findspot in Central Wales, The Museum of Wales online database in the archive Ancient Wales
0 notes
Text
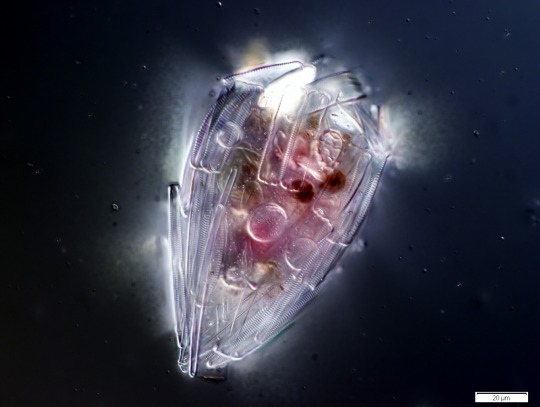
A testate amoeba which has covered itself with protective diatom frustules.
1 note
·
View note
Photo
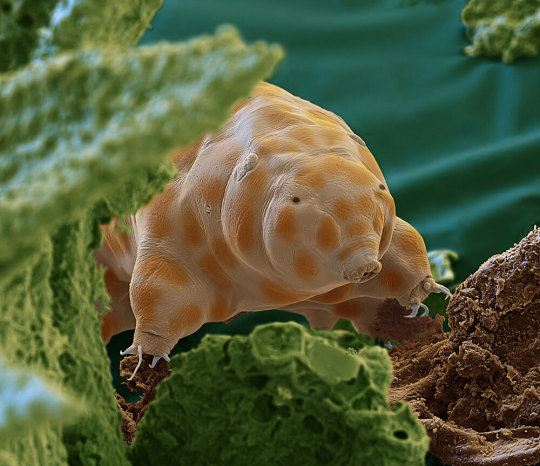
The newfound species seen here is one of about 1,300 known types of tardigrades. It was discovered in moss growing on dead tree trunks in Germany’s Black Forest. Far too small to see with the unaided eye, this creature is among billions of life-forms on the forest floor that are essential to the health of the planet. Magnified 2,400 times.
THESE TINY CREATURES ARE MARVELOUS
PHOTOGRAPH BY OLIVER MECKES AND NICOLE OTTOWA
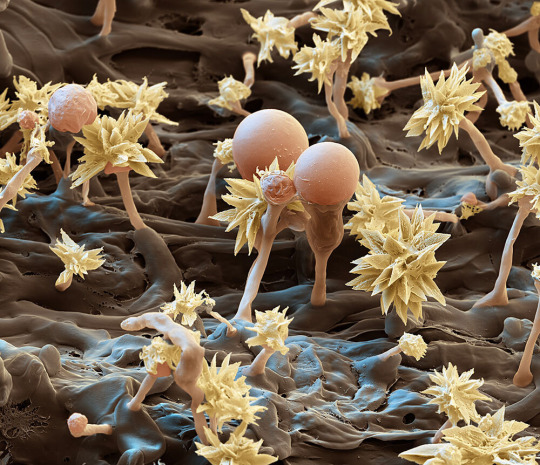
Fungi like this Resinicium bicolor—shown magnified 7,000 times—start breaking down dead trees by digesting lignin, the complex compound that helps form woody cell walls in plants.
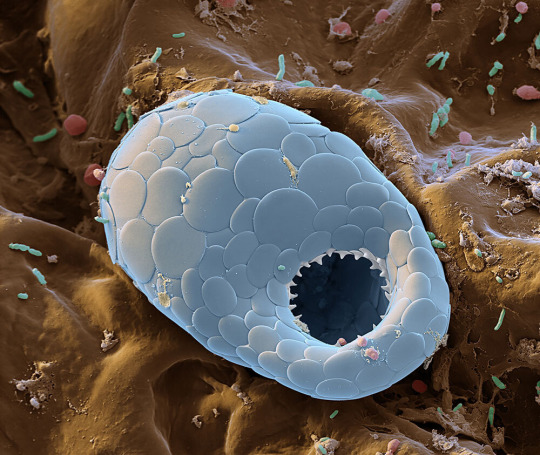
Scales of silica cover the single-celled body of a testate amoeba. These types of amoebas are named for the hard shells they create, possibly for protection against environmental changes within the forest litter.
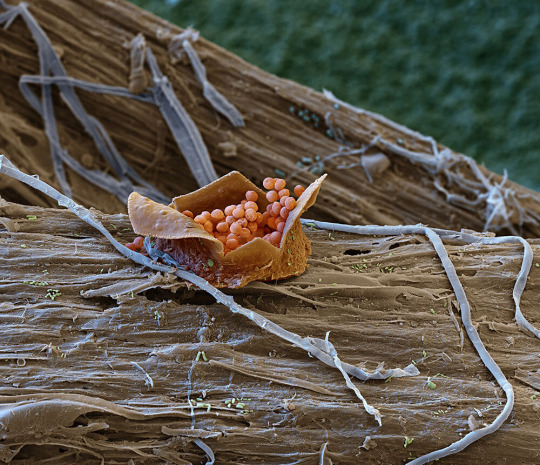
Resembling a fairy’s gift basket, the fruiting body of a slime mold, magnified 400 times, releases spores from its perch on woody debris draped in fungal filaments. Slime molds feast on other microbes found in decaying plant
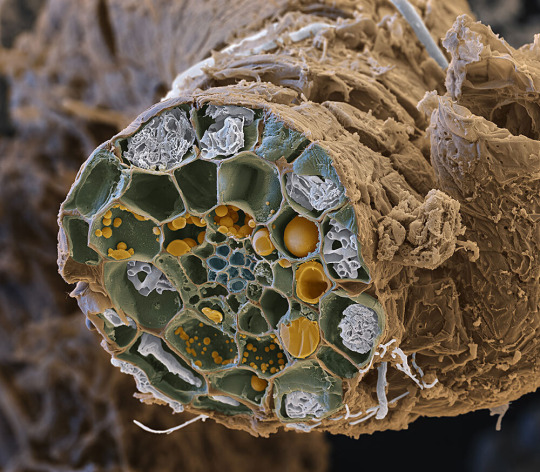
Some mycorrhizal fungi make their homes inside plant cells, as seen in this cross section of a European blueberry root. This allows soil residents of very different sizes to exchange nutrients—helping the forest
#oliver meckes#nicole ottowa#photographer#tardigrades#moss#germany#black forest#micro photography#nature#national geographic#fungi#resinicium bicolor#testate amoeba#slime mold#mycorrhizal fungi
644 notes
·
View notes
Text
@forseenconsequences This is exactly how I see it! Diatoms have a cell wall made out of silica and a very unique mechanism of creating it. They don’t have any other substances in their cell wall, the creatures that have calcium salts in their cells are testate amoebas, they are very different!
As for Bill I think his sides have a microscopic gap between two parts of his shell so that his limbs can come out. His limbs are 2D and have no bones, so it would make no problem.
I also think that the shell/exoskeleton is layered, like in insects which allows for limited flexibility. And he also has to have pigment cells somewhere because of his ability to change color. I can’t believe that I’m drawing diagrams, but:
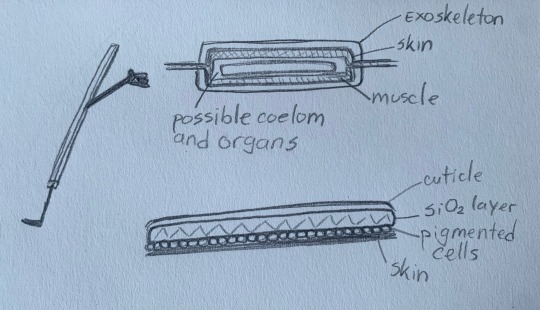
That thing on the left is Bill’s side profile with the gap where his limbs come out. Upper right is a cross section of his body in arm level. I thickened it up so that all structures can be seen more clearly. The bottom right image are the possible layers of his exoskeleton.
Weekly Test
(casual)







9K notes
·
View notes
Photo
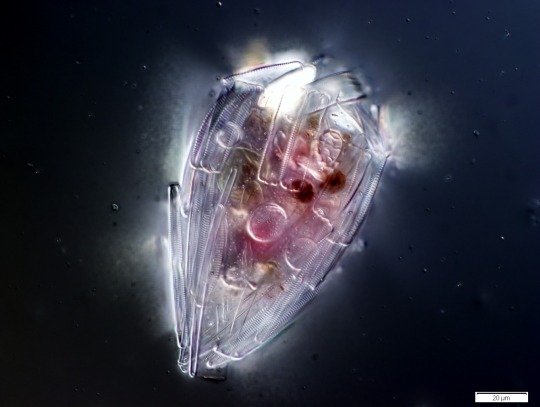
A testate amoeba which has covered itself with protective diatom frustules. Amoeba from the Collection Pénard MHNG. (via Wikipedia)
72 notes
·
View notes
Photo




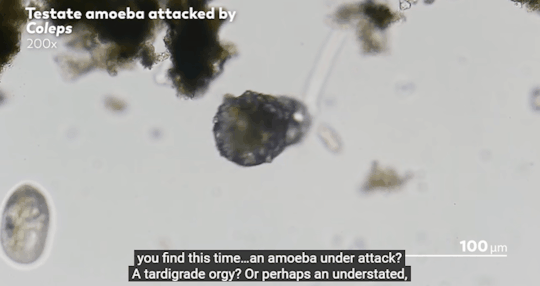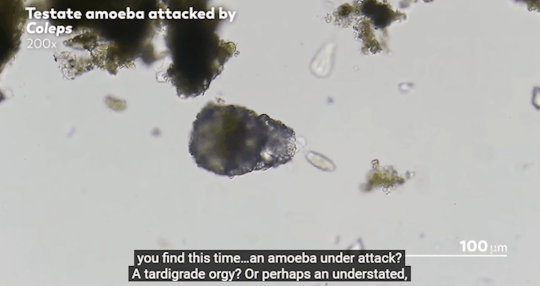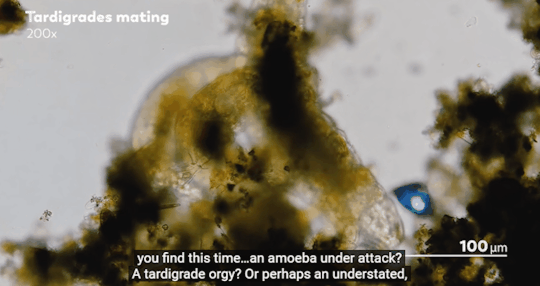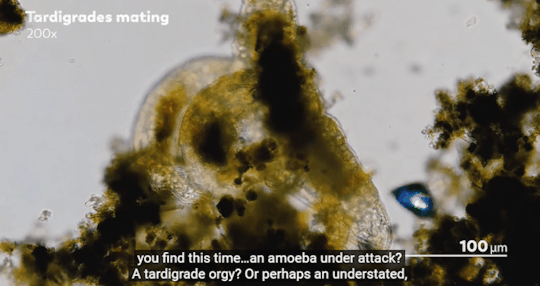

Journey to the Microcosmos- Lacrymaria: Vicious Long-Necked Predators
Images Originally captured by Jam’s Germs
Quote Spoken by Hank Green
#journey to the microcosmos#lacrymaria vicious long-necked predators#science#scienceblr#biology#bioblr#microbiology#microorganisms#microbes#lacrymaria olor#testate amoeba#coleps#tardigrade#oscillatoria#science aesthetic#science quote
202 notes
·
View notes
Text
・International Conference on Embodied Cognitive Science 2025(OIST, 沖縄, 2025年11月10日〜14日)にて口頭発表(Norihiko Kamaya, Katsunori Miyahara)を行います。 タイトル:Embodied agency at the cellular scale: a body-schema perspective on testate amoeba behaviors
1 note
·
View note
Text
Tree of Life 5: Unikonta (amoebae, fungi, and such)
Whew, this took a while! We continue our exploration of the diversity of life on Earth, branch by branch.
[Disclaimer: taxonomy is a complex, ever-changing field, and this overview is certainly not going to be exhaustive, especially concerning extinct groups]
← Part 4 (Angiospermae: flower plants) Part 6 (Metazoa: animals) →
Unikonta “one flagellum” is a rather diverse clade of Eukaryotes that group together many species of amoebae, slime molds, fungi, and animals. They (which is to say us) don’t share so many obvious features, but:
they are primarily heterotrophs (i.e. they take their carbon from organic sources), except for some species that entered symbiosis with algae; and
their cells have at most one flagellum, if any at all -- in contrast with flagellate algae, Euglenids, or Heterokonts, which typically have two flagella per cell (see parts 2 and 3 for more detail on those).
The group is more-or-less synonymous with Amorphea “shapeless” and Podiata “footed”, in reference to the fact that amoebae and animals have no rigid cell walls and can extrude appendages from some cells -- which is not true for fungi, though!
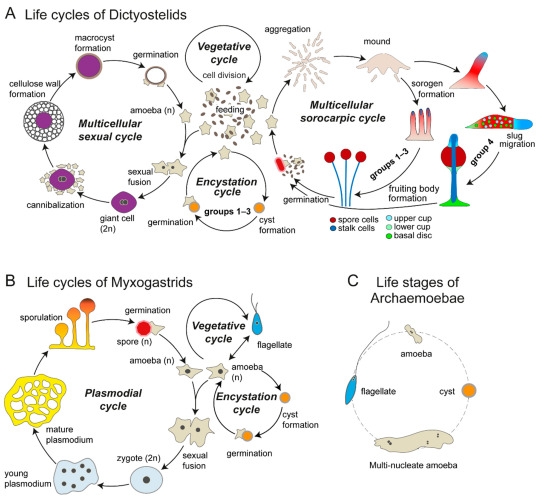
The different and wonderful life stages of amoebae -- I promise they will make sense soon. (Kin & Schaap 2021, “Evolution of Multicellular Complexity in The Dictyostelid Social Amoebas” (link))
1. Amoebozoa “changing animals”: The core clade of amoebas. (We have seen examples of amoebas, that is cells that can significantly alter their shape by extending and retracting cytoplasm-filled pseudopods, all over Eukarya -- see part 2.)
1a. Lobosa “lobed”: These amoebae have lost any trace of cilia or flagella, and move exclusively by pseudopods.
1a1. Tubulinea “tube-shaped”: Cells usually cylindrical in general shape, producing pseudopods that are themselves cylindrical and pushed by a central stream of granular cytoplasm. Amoeba dubia or Polychaos dubium is here, whose massively redundant genome is 200 times as large as ours (... maybe). Some Tubulinea, known as “testate amoebae” such as Arcella, are protected by a spherical shell of variable composition, with a single round opening from which pseudopos protrude.
1a2. Discosea “disk-shaped”: These amoebae are rounded and flattened; their pseudopods are small, thin, and contain only the transparent surface layer of cytoplasm. Here we found Acanthamoeba, which lives in soil and freshwater but can be an opportunistic parasite in humans.
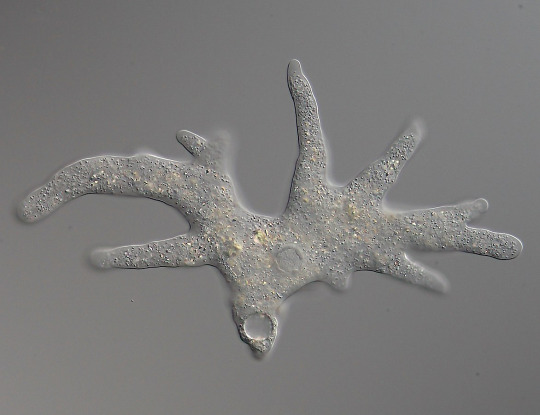
Amoeba proteus (Tubulinea), the most classical species of amoeba, extending its pseudopods. The whole thing is almost a millimeter across. (SmallRex, Wikimedia)
1b. Conosa “cone-shaped”: These ones may or may not have flagellate stages; if they do, the base of the flagellum has a cone-shaped microtubular basket aftre which the group is named.
1b1. Archaemoebae “ancient amoebae”: One of the old Archaezoa (see part 2), believed to be primitive Eukaryotes because they lack mitochondria. In fact, they just lost them as a consequence of living inside other organisms or such places where food is abundant and oxygen is not. Outside of hosts, they have to cease activity and withdraw into a cyst. They include Entamoeba histolitica, which causes amoebic dysentery in humans; the giant Pelomyxa palustris, which can measure several mm in length and carry thousands of nuclei; and the strange Mastigamoeba, which retains a flagellum even in its amoeboid stage.
1b2. Dictyostelida “net-like stalk”: One of the major groups of slime molds, very interesting organisms that alternate between unicellular and multicellular organization. Like the rather similar Acrasida, which are found very far away in the tree of life (see part 2), they are known as “cellular slime molds”. They grow as haploid unicellular amoebae, feeding by phagocytosis of bacteria and dead matter. However, when food becomes scarce, the separate cells emit a signal that causes them to join into a slug-like gelatinous body able to crawl around. This body eventually generates stalked fruiting bodies that scatter spores to the wind. The species Dictyostelium discoideum is a classical model in studies on cell communication and the evolution of multicellularity.
1b3. Myxogastria “slimy interior” or Myxomycota “slimy fungi”: The other major group of slime molds. In contrast with the “cellular” Dictyostelida, these ones are “plasmodial slime molds”: their cells don’t merely aggregate, but fuse together, forming a single mass of cytoplasm with thousands of unique nuclei, that is, a plasmodium. These can produce spores or flagellate “swarming” cells for dispersal, which, in turn, produce gametes for sexual reproduction. Fuligo septica (whose name means “infected soot”) and Physarum polycephalum are well known; the latter especially for its fantastic ability to self-organize its fluid body for efficient location and distribution of resources.
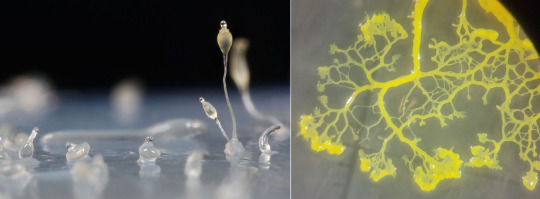
Two types of slime molds. Left: Dictyostelium discoideum (Dictyostelida), growing fruiting bodies from the slime. (Usman Bashir, Wikimedia) Right: Physarum polycephalum (Myxogastria), growing in a network to search food in its Petri dish. (Tim Tim, Wikimedia)
2. Opisthokonta “posterior flagllum”: So-called because their flagellate cells tend to push themselves forward with a flagellum at the back end of the cell, whereas flagellates in most other clades draw themselves forward with a flagellum beating at the front. (Think of sperm cells here; however, most animal cells are not flagellate, and most species of fungi have no flagella whatsoever.)
2a. Holomycota “all fungi”: The general neighborhood of Fungi; this clade is defined as the broadest possible clade that contains molds and mushrooms but not animals.
2a1. Nucleariida: A very small group of heterotrophic protists. It includes the genera Nuclearia, an amoeba with thin, thread-like pseudopods, and Fonticula, a cellular slime mold very similar to Dictyostelium.
2a2. Fungi: The kingdom of mushrooms, molds, and yeasts. They are all heterotrophic (unless you count lichens), extracting organic material from other organisms, usually as decomposers (indeed, they do most of the decomposition in all ecosystems), or sometimes parasites. They are never motile, except for a brief flagellate stage in some groups. Cells form thin filaments called hyphae, growing only at their tip. Hyphae contain multiple nuclei, and are surrounded by a cell wall made of chitin (a nitrogen-rich sugar polymer that also goes into the cuticle of insects). Like plants, they rely on this wall and water-filled vacuoles to keep their shape. They are usually haplontic (remember? That means the cells of their multicellular life stage have only one copy of each chromosome, rather than two as in animals). The earliest aquatic fungi appeared possibly over a billion years ago, and they started spreading on land around the same time as plants, in the Ordovician and Silurian periods (460-420 million years ago).
? 2a2a. Microsporidia: It’s unclear whether they are actual Fungi that reverted to unicellularity or just close relatives. They are intracellular parasites, which made them smallest among Eukaryotes, down to a single micrometer in length. They lost mitochondria and several other structures, but developed a harpoon-like polar filament which they use to penetrate into cells. They can produce tumor-like growths, called xenomas, in the organs of fish and Arthropods they infect.
2a2b. Blastocladiomycota “branching sprout fungi”: These basal groups still retain zoospores, that is, a unicellular flagellate stage of their lifecycle that allows easy dispersion in water. Blastoclads are microscopic, with one or more specialized cells forming at the end of the hyphae to produce spores, and root-like structures to absorb nutrients. They can be decomposers or parasites -- such as Physoderma maydis, which cause “brown spot disease” on maize leaves. Allomyces, like land plants (see part 3), has a haplodiplontic cycle, alternating a haploid body that produces gametes and a diploid body that produces flagellate spores.
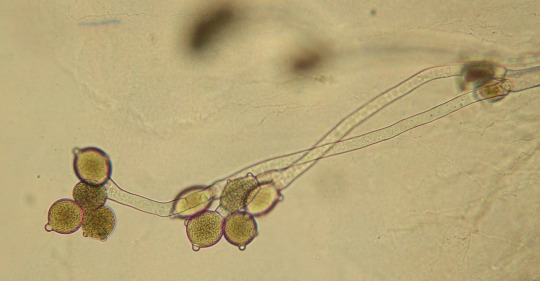
Allomyces sp. (Blastocladiomycota), with sporangia growing at the end of its hyphae. (TelosCricket, Wikimedia)
2a2c. Chytridiomycota “little pot fungi”: In practice, very similar to Blastocladiomycetes. Chytrids were found in the 400 million-years-old (Early Devonian) site of Rhynie Chert, which means they were on land among the very first plants. Batrachochytrium dendrobatidis is infamous as a scourge of frogs, to whom it causes potentially fatal skin infections, having already driven to extinction up to several dozens of species. The following groups of fungi have completely lost their flagella, and thus are grouped as Amastigomycota (“thread-less fungi”).
2a2d. Mucoromycota “slimy fungi”: The core of the old phylum Zygomycota “joined fungi”, now believed to be polyphyletic. Their hyphae don’t have internal partitions, so cytoplasm can flow freely along their whole length. The old name refers to the fact that they have upright, standing branches that can fuse their tip with their neighbors’ to form a diploid zygospore. This immediately undergoes meiosis to release haploid, non-flagellate spores. (Often this fusion can only occur between different mating types, which are not quite sexes because they are otherwise identical.) Mucoromycotans are known as “pin molds” because their erect hyphae topped with round sporangia make them look like pins stuck in their substrate. One such example is Mucor mucedo, which produces a white-grey fuzz on some cheeses; another is Rhizopus, which is used in Indonesia to ferment soy in making tempeh (R. oligosporus) and in Japan to make sake from rice (R. oryzae), and is commonly found on stale bread and old strawberries (R. stolonifer). Pilobolus crystallinus grows on dung piles, and has explosive vesicles that can shoot spores several meters away.

Closeup of a pin mold, identified as “probably Rhizopus stolonifer” (Mucoromycota), growing on a tomato. Note the upright white hyphae, each holding up a rounded sporangium. Most “fuzz” on moldy fruit looks like this. (Calimo, Wikimedia)
2a2e. Glomeromycota “yarn-ball fungi”: These fungi are also found as fossils in Rhynie Chert, and in a very important form: that of arbuscular mycorrhizae, which are structures formed by symbiotic fungi penetrating into the roots of a plant. The mycorrhizae depend on the plant for photosynthesis-produced sugar, and in exchange they provide the plant phosphorus and minerals. This association, starting from the very first terrestrial plants, is extremely important, and now found in as much as 80% of modern vascular plants (in many cases, the fungus belongs to genus Glomus). One exception is Geosiphon, which does not form mycorrhizae, but in compensation hosts a symbiotic Nostoc, a cyanobacterium you might remember from part 1, within its cells.
2a2f. Dikarya “two nuclei”: Most Fungi that you may have actually seen -- say, mushrooms -- are in this group. The name refers to a unique aspect of their reproduction: when haploid hyphae from different individuals, grown from spores, meet and meld, they exchange nuclei, as they would in any case of sexual reproduction. However, the nuclei do not merge, and the offspring will contain two nuclei in each cell. The nuclei merge into a diploid zygote only immediately before the meiosis that will produce new haploid spores -- but this occurs only within specialized structures that reside in a fruiting body. What is commonly called “mushroom” is simply the spore-producing fruiting body of a fungus, built of tightly-packed hyphae.
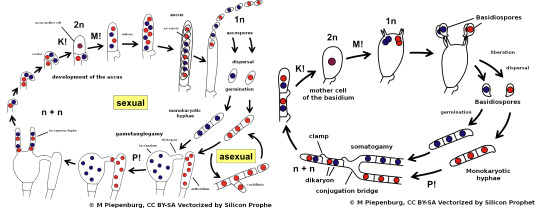
The generalized life cycles of Ascomycota (left) and Basidiomycota (right). Blue and red dots represent cell nuclei from different parents. “n”/”1n” and “2n” represent respectively haploid and diploid cells. “M!”, “P!”, and “K!” represent, respectively, meiosis (halving of genetic material), plasmogamy (fusion of the cytoplasm of two cells), and karyogamy (fusion of two nuclei). In plants and animals, plasmogamy and karyogamy occur almost at the same time during fertilization, but in Fungi they’re distinct events. (Silicon Prophet, Wikimedia)
2a2f1. Ascomycota: This large group (three quarters of all known species of Fungi) is named after its peculiar reproductive organ, the ascus (“sack”). This is a single rod-shaped cell, at the end of a hypha, in which the two parent nuclei merged and then underwent repeated meiosis to become (usually) eight unique haploid nuclei. When each nucleus has built a spore around itself, the ascus will break open or dissolve to release the spores.
2a2f1a. Taphrinomycotina: This early-diverging group does not show obvious similarities. Most examples are microscopic: Schizosaccharomyces pombe is a type of yeast, which is to say a fungus that has returned to a unicellular form and no longer grows into hyphae (though it still produces spores by turning into an ascus with four nuclei); found often in fruit and alcoholic drinks, it can also reproduce by binary cell division. Pneumocystis jirovecii is also unicellular, and a parasite in human lungs, where it causes pneumonia. The namesake genus Taphrina, which causes leaf curl disease in peach and almond trees, alternates a yeast-like form and hyphae, finally forming a layer of asci on infected leaves. One exception is genus Neolecta, which forms smooth, lobed fruiting bodies that are bright yellow and a few cm tall.
2a2f1b. Saccharomycotina “sugar fungi” are a group dominated by yeasts. They, too, become a four-spore ascus after meiosis; but before that, when they are still diploid thanks to the fusion of nuclei, they can reproduce by asymmetrical budding, which leaves a visible scar on the mother cell. Most important for us is the brewer’s yeast Saccharomyces cerevisiae, whose fermentation of sugar produces both ethyl alcohol and carbon dioxide, and is therefore used to brew beer and leaven bread. Another one is Candida albicans, which is commonly found in the human mouth and vagina but can become a pathogen when our immunitary system falters.

A few cells of the baker’s or brewer’s yeast, Saccharomyces cerevisiae (Saccharomycotina), producing daughter cells by budding. Yeasts are unicellular descendants of multicellular ancestors. (Masur, Wikimedia)
2a2f1c. Pezizomycotina: The fruiting body (ascocarp) is composed of one or more bowl- or flask-shaped apothecia, each with asci coating its concave inner surface. This group counts fourteen classes, highly variable. A few examples:
Pezizomycetes contains most Ascomycotan mushrooms. “Cap fungi” like Aleuria and Sarcoscypha have flattened reddish ascocarps that look like discarded orange peels. The edible morels (Morchella) have a sponge-like cap, formed by many hollow apothecia clustered together. The highly-prized truffles (Tuber) grow underground as root symbionts of oaks and other trees.

A group of common morels (Morchella esculenta, Pezizomycetes) in a forest in Illinois. Each “hole” in its cap is an apothecium internally coated with spore-bearing asci. (DrewHeath, Wikimedia)
Other classes are dominated by molds. Sordariomycetes is home to the bread mold Neurospora crassa, an important model organism in genetics; Fusarium venenatum, cultivated as meat substitute under the name of Quorn; the ergot Claviceps purpurea, which covers infected rye with black horn-like grotwhs, and does dreadful things to people who consume it. Eurotiomycetes has Penicillium, which is used in cheesemaking and from which the first antibiotic (penicillin) was extracted, and Aspergillus, a toxic black mold that can infest damp houses. These molds often have entirely asexual life cycles, and produce huge numbers of spores with no need of sexual recombination (the sheer amount of spores they release is, indeed, the reason they can be dangerous).
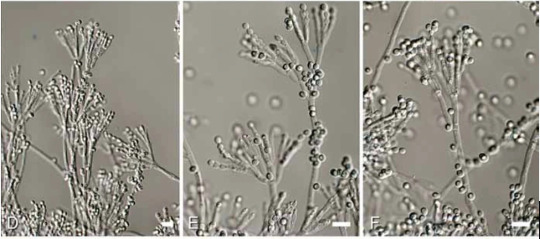
The microscopic fronds of Penicillium rubens (Eurotiomycetes), to which we owe the discovery of antibiotics. Such molds grow as blue-greyish velvet on old fruit. (Houbraken & al. 2011, “Fleming’s penicillin producing strain is not Penicillium chrysogenum but P. rubens.”)
Lecanoromycetes contains the majority of lichens. These fungi have formed a symbiotic relationship with unicellular algae or cyanobacteria that live inside their tangle of hyphae, just close enough to the surface to receive light. The fungi produce rock-dissolving acids to extract minerals, passing them to the algae which provide their host with sugars; this combination allows lichens to survive even in the most barren environments. They range in shape from thin layers encrusting rocks, to bush-like structures such as the reindeer lichen (Cladonia rangiferina) or the Iceland moss (Cetraria islandica).
Finally, let me mention Arthrobotrys, from the Orbiliomycetes: this is a rare case of a carnivorous fungus, whose hyphae form microscopic loops to trap and digest Nematode worms.
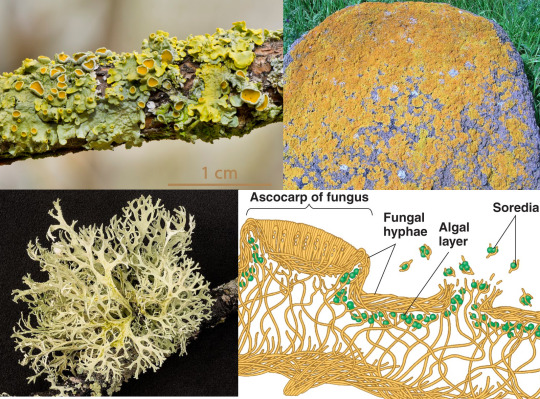
Top left: a foliose (leafy) lichen, Xanthoria parietina, growing on a branch; the little cups are the ascocarps. (Norbert Nagel, Wikimedia) Top right: a crustose (crusty) lichen, Caloplaca marina. (Roger Griffith, Wikimedia) Bottom left: a fruticose (bushy) lichen, Cladonia portentosa. (Hubertl, Wikimedia). Bottom right: the microscopic structure of a generic lichen. (Benjamin Cummings, Pearson Education; this is the closest link I could find)
2a2f2. Basidiomycota: Again, this group is named after its reproductive structure, the basidium (”little pedestal”). Again, the basidium is a single club-shaped cell in which the parent nuclei merge before undergoing meiosis. The resulting unique haploid nuclei emerge at the end of the basidium as (usually) four spores that are (usually) forcibly expelled upon maturity. These fungi have septa within their hyphae which keeps the nuclei separated from each other.
2a2f2a. Ustilaginomycotina: A clade of parasites of vascular plants, generally known as “smuts”. They start their life as haploid, saprophytic yeasts, which merge two by two in presence of host plants to form parasitic hyphae. These hyphae end in appressoria, specialized invasive structure that can pierce through any tissue thanks to hydraulic pressure. The best known species is the corn smut (Ustilago maydis), which forms (edible!) grey "mushrooms” on infected maize ears.
2a2f2b. Pucciniomycotina or Urediniomycotina: Most species of this clade are also plant parasites: they cause stunted growth and yellowing of leaves, as well as rough yellow-to-red growth which gave them the common name of “rusts”. Puccinia graminis is one of the most important pathogens of wheat and barley, while Helicobasidium longisporum attacks plants as diverse as apples, potatoes, and soy.

Left: corn smut (Ustilago maydis, Ustilaginomycotina) causes infected kernels to swell into grey “fruits” filled with the fungus’ spores; under the name huitlacoche, they are eaten in Mexico. (Kai Hirdes, Wikimedia) Right: A pear leaf infested by the pear rust (Gymnosporangium sabinae, Pucciniomycotina); the black blisters release airborne spores. (Maurice Flesier, Wikimedia)
2a2f2c. Agaricomycotina: And here we find Basidiomycotan mushrooms. Their fruiting body (basidiocarp) often has the distinctive umbrella-like shape with cap and stalk, and has lamellae coated with the basidia.
Two of its subdivisions, Tremellomycetes and Dacrymycetes, comprise the so-called “jelly fungi”, whose fruiting body is indeed gelatinous or rubbery: from the first group, the witches’ butter (Tremella mesenterica), which grows as a parasite on the branches of broadleaf trees; from the second, the yellow stagshorn (Calocera viscosa), which forms antler-like growths on the ground. They are usually edible, but not very appreciated for taste.
Agaricomycetes contains a vast diversity of mushrooms, including many edible ones: the common champignon (Agaricus bisporus) and shiitake (Lentinula edodes), which have distinctive spore-bearing "gills” and are the most consumed mushrooms in the world; the trumpet-shaped golden chanterelle (Cantharellus cibarius); the club-shaped puffball (Lycoperdon perlatus), which blows clouds of spores out of a hole at its top; the porcino (Boletus edulis), whose spores must instead escape through pores on the cap’s underside. Many others of course are toxic, or have hallucinogenic properties, such as the red-and-white Amanita muscaria or the many species of Psilocybe. The skinhorn (Phallus impudicus) is famous both for its unique phallic shape and for stinking of rotten meat -- as it should, since it relies on carrion flies to spread its spores. Polyporales, such as Ganoderma, include those flat, disk-shaped mushrooms that grow on the side of living or dead tree-trunks. A particular colony of Armillaria ostoyae in a forest in Oregon spans nine square km, which makes it one of the largest living organisms on Earth if it’s indeed a single mycelium.

A selection of Agaricomycetes. Top left: the “earthstar” Geastrum triplex. (Josef F. Stuefer, Wikimedia) Top right: a bracket fungus (Ganoderma applanatum) growing on an old trunk. (Eric Steinert, Wikimedia). Bottom left: golden chanterelles (Cantharellus cibarius). (Andreas Kunze, Wikimedia) Bottom right: the psychedelic Psilocybe semilanceata. (Arp, Wikimedia)
2b. Holozoa “all animals”: The mirror image of Holomycota, defined as th broadest possible clade that includes animals but not Fungi.
2b1. Ichthyosporea “fish spores”: All unicellular, with a chitinous cell wall; some flagellate, other amoeboids. They are mostly parasites of animals, especially amphibians and fish (most importantly, Dermocystidium, which lives in the skin and gills of freshwater fishes).
2b2. Choanoflagellatea “funnel flagellates”: Now these are interesting. These tiny protists all have a flagellum, around which stands a ring of rigid microvilli (effectively immobile cilia) that form a sort of cone-shaped funnel, or collar. When swimming, the flagellum generates a current of water away from the cell, which pushes it forward, like sperm cells; but it also creates a current of water into the collar and out of its central opening, so that smaller cells and other particles of food are trapped between the microvilli and may be ingested. Often they are surrounded by a glycoprotein gelatin, that might attach the cell to the seafloor (as in Codonosiga), to each other (as in Salpingoeca), or both (as in Proterospongia). Molecular studies suggest that Choanoflagellates are the closest kin to Animals; as it happens, sponges have some very choanoflagellate-like cells (see part 6 on that).
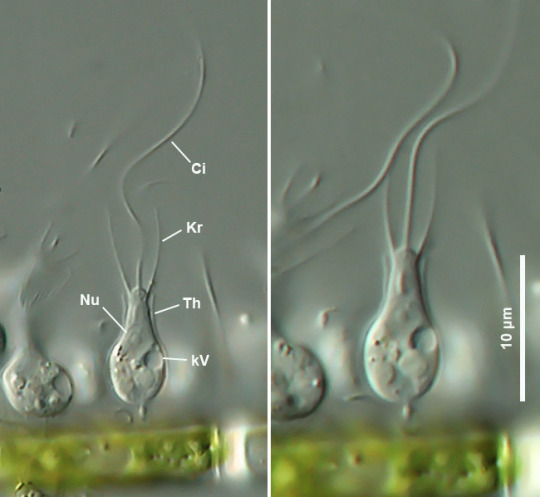
Salpingoeca (Choanoflagellatea) attached to a green alga, showing both the flagellum (Ci) and the sides of the collar (Kr). Water is drawn into the collar from the sides and expelled upward thanks to the motion of the flagellum, trapping food between the microvilli. (Ole Riemann, Mikro-Forum)
2b3. Metazoa or Animalia: heterotrophic and multicellular, with at at least two distinct layers of tissue: all animals from sponges to hummingbirds and from slugs to mosquitoes are here, including ourselves. See part 6, and all the following.
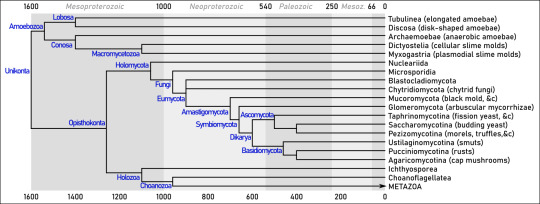
Summary. Dates (mostly from Parfrey &al 2011 and Tedersoo &al 2018) are in millions of years ago.
Sources Adl & al (2019), “Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes” (link) Brown (2014), Principles of Microbial Diversity, APM Press Brusca (2016), Invertebrates (3rd edition), Sinauer Cavalier-Smith & al (2015), “Multigene phylogeny resolves deep branching of Amoebozoa” (link) Deacon (2005), Fungal Biology (4th edition), Blackwell Derelle & al (2015), “Bacterial proteins pinpoint a single eukaryotic root” (link) He & al (2014), “An Alternative Root for the Eukaryote Tree of Life” (link) Hickman & al (2008), Integrated Principles of Zoology (14th edition), McGraw-Hill Parfrey & al (2011), “Estimating the timing of early eukaryotic diversification with multigene molecular clocks“ (link) Schilde & Schaap (2013), “The Amoebozoa” (link) Tedersoo &al (2018), “High-level classification of the Fungi and a tool for evolutionary ecological analyses“ (link)
20 notes
·
View notes