#eutriconodonts
Text
Volant Vertebrates of Foliest
Part One - Therodactyls
Fairies share the skies of Foliest with a range of other aerial organisms. Insects and spiders have both evolved powered flight, while certain rainforest worms have developed the ability to glide between trees. Among vertebrates, flight has evolved thrice. The earliest to take to the skies were the jimmylegs, descended from ancestors much like Sharovipteryx, which adapted their hindlimbs into wings. The next were the birdonts, basal archosaurs that glided on outstretched ribs, which were then modified until capable of powered flight. Finally, the mammals produced their volant representatives, the therodactyls.
Evolving in Foliest's late Jurassic from gliding, eutriconodont ancestors much like Volaticotherium, they got their start as insect hunters with some larger, coastal forms specialized in fish. Floral turnover in the Cretaceous resulted in the expansion of closed-canopy forests, and this environment seems to have favoured the therodactyls over the birdonts, in many ways their ecological rivals. They underwent rapid bursts of evolutionary radiation in this period and slotted into diverse niches they had never held before. By the middle Cretaceous, the two clades had reached an equilibrium. Therodactyls tend to be the largest flying vertebrates in their ecosystems and dominate coastal, wetland and forest canopy habitats, with birdonts more numerous in plains, scrublands and the remaining open forests. Jimmylegs, meanwhile, avoid competion simply by staying small and mostly nocturnal.

Pictured here are some representative species. At the top is a macrocarnivore that primarily hunts other therodactyls on the wing. Middle is a plunge-diving piscivore that soars above the waves and pursues fish for short distances underwater. Bottom left is a nut and seed eater that has convergently evolved traits similar to rodents, such as cheek pouches and continuously growing incisors. Bottom right is a frugivore that encourages algae and bryophytes to colonize its fur, providing camouflage against predators that ambush from above (easier than on Earth thanks to the rate at which fairy-tended plants grow).

One of their more unusual traits is the fact that they utilise all four limbs for flight, similar to certain dinosaurs such as Microraptor. With a wing membrane stretched between digits, as with bats, they can alter the shape and size of the wing by flexing certain fingers and toes. This sequence demonstrates a predatory therodactyl in three modes of flight. The first is the typical wing position, with a single, large surface on each side allowing efficient soaring. The second shows the longest digit on each limb being folded, bunching up the membrane and separating each wing into two, improving maneuverability at the cost of greater energy expenditure. The third portrays all digits on the feet being tucked close to the body, reducing the animal to a single, smaller set of forewings. When hunting, a therodactyl would begin in the first position while locating prey, switch to the second if the initial strike failed and a chase was required, and the third when the hindwings were needed not for flight, but to strike with the talons.
#speculative evolution#speculative biology#speculative zoology#spec evo#spec bio#spec zoo#prehistoric mammals#eutriconodonts#volaticotherium#therodactyls#worldbuilding#art#creature art#creature design#fauna of foliest#fairies
2 notes
·
View notes
Text


Celebrating bat appreciation week with the rather non-bat volaticothere commissions of @paleoart, mammals that flew 100 million years before bats did!
#volaticothere#volaticotheria#mammal#eutriconodont#eutriconodonta#paleontology#palaeontology#paleoblr#palaeoblr
134 notes
·
View notes
Text

Sleepy Repenomamus and awake Mei
27 notes
·
View notes

Text

Although most Mesozoic mammals were rather small, a few different lineages produced some pretty hefty-sized forms – most notably the metatherian Didelphodon, the gondwantherians Adalatherium and Vintana, and the eutriconodont Repenomamus.
And now we've got another one to add to that list.
Patagomaia chainko lived towards the end of the Cretaceous, about 70 million years ago, in what is now Patagonia near the southern tip of South America. Known from some partial leg and hip bones, it was potentially the largest known Mesozoic mammal yet discovered – estimated to have been similar in size to a modern bobcat, roughly 50cm tall at the shoulder (~1'8") and weighing around 14kg (~31lbs).
Distinctive anatomical features of the bones indicate it was an early therian mammal, the group that contains both modern marsupials and placentals, but it can't currently be classified any more specifically than that. Mesozoic therian fossils are very rare in the southern continents, so Patagomaia's presence in late Cretaceous South America adds to their known range and diversity, as well as providing an example of surprisingly large body size for the time.
Without more material it's impossible to tell what Patagomaia's ecology was. I've gone for a fairly generic life appearance here, and while what's known of its joints and muscle attachments doesn't indicate climbing specializations, plenty of unexpected tetrapods still like to get up on tree branches.
———
NixIllustration.com | Tumblr | Patreon
#science illustration#paleontology#paleoart#palaeoblr#patagomaia#theria#mammal#mesozoic mammal#art#big mesozoic mama#truly one of the most mammals of all time
338 notes
·
View notes
Text
i think it´s FINALLY time to do this,
alot of idea´s have come up from
Gurren Lagann
Ice age 3
Suria
the Lost world
atlantis the Lost Empire
and alot of other things too, and there might even be a buck like character
unsure if wanna make him a Pine marten, Eutriconodont, Didelphodon even a honey badger

still tho got a cool idea.
0 notes
Photo

So here is my V2 of my old picture of the species Ichthyoconodon jaworowskorum, an unsual eutriconodont found in the sea deposits of the lower cretaceous Morroco, being its only known remains formed by two lower jaw Molars, MNHN SA 46 (a) and SA 78 (b).
Is really little of what it is known about this as with two teeth there isn't much what can be inferred, just probably that they were some inhabitants of the coastal region of Northwest coast of africa... and that's pretty much it, nothing else can be taken from 2 isolated teeth.
Honestly is a shame that this and many relatives of these are just little know from few complete specimens, and just mostly being jaws and teeth what it is found.
10 notes
·
View notes
Text
MESOZOIC MONTHLY: Volaticotherium
by Lindsay Kastroll
Once again, spring has sprung. Prepare to see the gorgeous forests of Pennsylvania launch back into action. I, for one, can’t wait to get outside and explore as the weather continues to improve. I was recently reminded of the fact that Pennsylvania is home to two species of flying squirrels, and I am definitely adding them to my list of things to see. But of course, this is Mesozoic Monthly, so flying squirrels can’t be the stars of this article. Instead, the superficially flying squirrel-like “ancient gliding beast” Volaticotherium antiquum is stealing the spotlight!
Although Volaticotherium was about the size of a modern flying squirrel at 5–6 inches (13–15 cm) long, it belonged to a group of early mammals called eutriconodonts that includes some of the largest mammals that lived alongside non-avian dinosaurs. “Eutriconodont” means “true three-coned tooth,” in reference to the three longitudinally aligned cusps on their molars. Although not all mammals today have three-cusped molars, the ancestors of modern mammals did. Does this mean that modern mammals evolved from a eutriconodont? The answer is no, though they did evolve from a mammal with eutriconodont-like teeth.
We can split modern mammals into two main groups: the monotremes, which are egg-laying mammals like the platypus, and the therians, which include both marsupial and placental mammals (like kangaroos or humans, respectively). The ancestors of monotremes diverged (meaning, formed their own ‘branch’ of the evolutionary tree) before eutriconodonts and therians evolved. Eutriconodonts and therians share a different, more recent, and as-yet unknown common ancestor. Monotremes, therians, and eutriconodonts actually lived alongside one another for over one hundred million years before eutriconodonts became extinct near the end of the Cretaceous Period (the third and final division in the Mesozoic Era, or ‘Age of Dinosaurs’).

This flowchart represents a simplified phylogeny (aka, evolutionary tree) of the relationships discussed in the previous paragraph. A lot of ‘branches’ and intermediate steps are missing from this phylogeny to make it easier to follow.
The canines and molars of eutriconodonts were pointy, suggesting that these mammals were carnivores or insectivores. Volaticotherium is no exception, which makes it particularly unique, as most other gliding mammals are herbivores! Because it was so small, Volaticotherium was probably an insectivore, but a larger cousin, Jugulator, could probably eat small vertebrates. As an arboreal glider, Volaticotherium could soar from tree to tree to catch insects in midair. Instead of wings, it had a patagium, a broad flap of skin that stretched between the fore- and hind limbs, creating enough surface area to achieve gliding descents. The various limb adaptations necessary to make Volaticotherium an efficient glider also made it poor at maneuvering on the ground. It can be hard to understand why an animal would evolve features that would hinder its terrestrial movement, and multiple hypotheses have been put forth to try to explain this. Most of these focus on the benefits of leaping out of trees to escape predators or to quickly traverse territory between arboreal food sources, scenarios based on herbivorous mammals. Because Volaticotherium was a gliding predator, perhaps gliding conferred other advantages to this eutriconodont.

Restoration of Volaticotherium in mid-glide by Jose Antonio Peñas, used with permission. Take note of those sharp canine teeth, useful for catching tasty insects! You can find more of Peñas’ art on their DeviantArt, ArtStation, or YouTube.
The fossilized remains of Volaticotherium were found in a layer of rock called the Daohugou Bed in China. This deposit consists of lakebed sediment and volcanic ash compacted into solid rock over millions of years as more heavy sediment was deposited on top of it. There is a debate about how old the Daohugou Bed is, but most estimates place it near the middle or end of the Jurassic Period (the middle period of the Mesozoic). Getting the timing right is important. Because Volaticotherium is among the oldest known gliding mammals, its discovery pushes the origin of mammalian gliding back as much as 70 million years earlier than previously thought!
A variety of factors have led geologists to struggle in determining the age of the Daohugou Bed. In an ideal geologic record, rock layers would be perfectly horizontal, creating a continuous stack with the oldest layers on the bottom and the newest layers on top. However, this is rarely the case. Sediment may be eroded before new layers are deposited, creating a gap of time without record in that sequence of rocks. This phenomenon, where two rock layers do not represent a continuous progression of time and have a gap of data missing between them, is called an unconformity. Other issues with dating rock layers involve the squeezing, stretching, folding, melting, and chemical alteration of rock layers when they’re subjected to geologic processes. These forces can result in old rock layers being placed on top of younger ones, making it hard to determine the actual sequential order of the rocks. Changes can also occur within the minerals that compose the rocks, making radiometric dating much more difficult.
The Daohugou Bed has an unconformity above and below it, and it has been folded, which makes attributing an exact age to it that much harder. When you go out hiking in the beautiful spring weather on the horizon, take a moment to look at the rock outcrops you pass and think about what those layers might have experienced on their journey to where they are today. And if you continue your hike after sunset, be sure to keep your eyes peeled. If you’re lucky, you might just catch a glimpse of a flying squirrel gliding through the forest!
Lindsay Kastroll is a volunteer and paleontology student working in the Section of Vertebrate Paleontology at Carnegie Museum of Natural History. Museum staff, volunteers, and interns are encouraged to blog about their unique experiences and knowledge gained from working at the museum.
23 notes
·
View notes
Text
Barosaurus lentus

By Ripley Cook
Etymology: Heavy Reptile
First Described By: Marsh, 1890
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria, Plateosauria, Massopoda, Sauropodiformes, Anchisauria, Sauropoda, Gravisauria, Eusauropoda, Neosauropoda, Diplodocoidea, Diplodocimorpha, Flagellicaudata, Diplodocidae, Diplodocinae
Status: Extinct
Time and Place: Between 150 and 149 million years ago, in the Tithonian of the Late Jurassic

Barosaurus is known from the Brush Basin Member of the Morrison Formation in South Dakota, Utah, and Wyoming. Potential specimens of Barosaurus are known from other locations of the Morrison Formation; the entire range of this habitat at the time of Barosaurus is shown below in green (with the range of Barosaurus inside of it, in blue).

Physical Description: Barosaurus in a lot of ways is a fairly typical Diplodocid sauropod - long, large, and with a distinctive whip-tail. But, when you dig under the surface, Barosaurus has nothing truly “typical” about it. The neck of Barosaurus is next-level in its length - and the tail is ridiculous to match. In fact, the estimates of the length of Barosaurus get huge - it was probably more than twenty-six meters long, and some of the most upper estimates of Barosaurus have it at fifty meters long! This would make it one of, if not the, largest known dinosaurs - and certainly the longest! Though it does have a long tail, it differs in appearance from its cousin Diplodocus primarily by having a proportionally longer neck and shorter tail. It was also more slender than Apatosaurus, though it was longer than that contemporary. How did Barosaurus get such a long neck? It literally converted one of the back vertebrae into a neck vertebra! This is so fascinating that I can’t get over it - its close relatives, like Diplodocus, did not employ this to get a longer neck, indicating Barosaurus was using its long neck for things that its cousins were not. Barosaurus was also weird in not having as high of spines on its vertebrae as its cousin Diplodocus and other members of the group. In addition to all of that - it had shorter vertebrae in the tail, which made it shorter than in other members of this group! Interestingly, the bones on the underside of the tail were forked and had forward spikes, which would have given it similar strength to that of Diplodocus; it was probably still a whiptail like other members of this group, though not as much of one as its relatives.

By Slate Weasel, in the Public Domain
Of course, the distinctiveness of Barosaurus is primarily limited to the length of the animal and its spine. In terms of limbs, it had fairly identical limbs to its cousin Diplodocus, though it did have fairly long forelimbs compared to its cousin (by… an almost imperceptible amount, however). Though the feet of Barosaurus aren’t known, it is reasonable to suppose that it would have had feet similar to Diplodocus - with only one claw on the front feet and three small claws on the hind feet. The skull of Barosaurus is not known, but it probably would have been long and low, with peg-like teeth in the front of the jaws for grazing on plants. Its neck was not very flexible in the vertical sense, but it was much more flexible in sweeping from side to side. It is possible that there were spikes of some sort at the end of the tail, which would have packed quite a punch when the tail was used to whip other animals. And, finally, it would have been entirely - if not almost entirely - scaly all over its body. It is also possible that Barosaurus may have featured some brilliant colors, especially in the tail, for communication with other members of the species.
Diet: Barosaurus would have primarily fed on high-level vegetation, able to reach much of it at its natural neck height and then - on top of that - being able to rear up to 50 meters high via going on its hind legs. However, a lack of vertical reach in terms of neck flexibility means that it probably would have swept over a wide area for food, rather than going up and down in the tree level like other Diplodocids. This would have allowed Barosaurus to move very little - if at all - while eating, instead of moving over large distances in search of vegetation.

By Scott Reid
Behavior: Barosaurus was not especially common in its environment, so the question of its social nature is actually somewhat important. Fossil evidence indicates at least some sociality in other Diplodocids - herding, or at least small herds, of other sauropods on the Morrison are clear from fossil evidence and trackways. The question remains - did Barosaurus do what its cousins did? The question is, of course, up in the air without more fossil evidence. It is possible that, in an environment with hundreds and hundreds of large sauropods to feed, Barosaurus may have been more solitary to aid in getting enough food without competing too much with one another. Alternatively, it may have also lived in social groups, allowing for the safety of weaker members of the herd and more cohesiveness in finding food.
Barosaurus, like other Diplodocids, would have been able to rear up on its hind legs to get food. This action would have also made Barosaurus even taller than usual, which would have been fairly imposing to predators nearby. It had a whip-tail, which would have allowed Barosaurus to make very loud sonic cracks in the air; if that tail was covered with spikes, as in other members of the group, it would have also lacerated the skin of other dinosaurs. Still, even without spikes, it would have packed quite a punch for any predators that might have tried to attack it. The sounds of the tail would have been a warning; it is possible that such sounds would have been used in communication with one another, and potentially even display in competition for mates and food and similar things. The impossibly long neck probably was also a sort of sexual display structure, since the longer neck indicated being able to reach more food without walking around. It is uncertain whether or not it would have taken care of its young; while there is no evidence either way - which usually would lead to concluding it did, given the fact most living archosaurs do and there’s extensive evidence of such in extinct dinosaurs - other sauropods (aka the titanosaurs) probably didn’t. So, for now, the jury on that is out.

By Fred Wierum, CC BY-SA 4.0
Ecosystem: Barosaurus lived in the Morrison Formation - an extensive, expansive semi-arid, seasonal floodplain that covered most of Western North America during the Jurassic and was filled with iconic dinosaurs and other animals that we usually think of when we think of the “Jurassic.” Though the Morrison was as arid and open as a modern savanna, the lack of extensive flowering plants at this time rendered the habitat more like a ridiculously huge scrubland. There were a variety of trees - conifers, ginkgos, cycads, and tree ferns - dispersed among the bushes and horsetails and other plants. They congregated around rivers, which were havens of life amongst the arid territory. At the time of the Brushy Basin Environment - the last part of the formation, where Barosaurus could be found - this environment was much muddier and wetter, potentially indicating a change in ecology that would lead to the end of the Morrison Formation, and an extinction of the animals there. There were also expansive volcanic explosions that lead to much of the preservation we see there. A large salt lake present would have been a major feature of the environment, and it was connected to extensive wetlands that formed a break in the wider scrubland around the habitat.

By Danny Cicchetti, CC BY-SA 3.0
Barosaurus may be known from the entire Brushy Basin Environment of the Morrison; however, confirmed fossils of this dinosaur are only known from a few sites. So, in my map above, I give two colors - the wider green color to show the whole ecosystem, aka the wider area that Barosaurus may have ventured in to; and the smaller blue color to show the confirmed range of this dinosaur. In that confirmed range, Barosaurus lived alongside a lot of other animals - in fact, there’s a reason the Morrison is so iconic - its characteristic and distinctive fossils, both of dinosaurs and not of dinosaurs. Barosaurus has been found in, literally, the same sites as other animals - it is known to have lived alongside the predator Allosaurus; in another site, turtles and Pseudosuchians and the Choristodere Cteniogenys, as well as Allosaurus and the more bulky sauropod Camarasaurus; in yet another, Barosaurus lived alongside many turtles, the Pseudosuchians Hoplosuchus and Goniopholis, the tuatara-like Opisthias, and a wide variety of dinosaurs - other sauropods like Diplodocus Apatosaurus and Camarasaurus, predators like Allosaurus Torvosaurus and Ceratosaurus, and Ornithiscians like Stegosaurus Dryosaurus and Uteodon. So, Barosaurus was a part of a very wide and diverse community, with a great diversity in terms of herbivores and predators that would have attacked Barosaurus. That being said, there were many other animals that may have lived alongside Barosaurus, based on just… probability, even though they weren’t found directly with it. There were other stegosaurs like Alcovasaurus and Hesperosaurus; more small running herbivores like Nanosaurus; larger bulky bipedal herbivores like Camptosaurus; more sauropods, including Apatosaurus and Supersaurus; and smaller predators that would have probably been more of a threat to Barosaurus young than adults - Marshosaurus, Coelurus, Ornitholestes, and Stokesosaurus. Sadly, the organization of the Morrison is something of a mess - so, while many other dinosaurs and animals lived alongside Barosaurus, we can’t exactly be sure which ones. There were probably a variety of Multituberculate, Tinodontid, Eutriconodont, and Dryolestoid mammals, as well as others; some pterosaurs were probably there like Harpactognathus, and, of course, there were amphibians as well. This makes the Morrison one of the better examples of an environment to highlight as a representative of a particular time in Earth’s history - since it showcases so many different living things!

By José Carlos Cortés
Other: Barosaurus was a close relative of Diplodocus, though it is difficult to determine how close, as the evolutionary relationships between the Diplodocids are still being worked out via phylogenetic studies. It is possible that an offshoot of Diplodocus (which were around before Barosaurus evolved) split to take advantage of not moving much to eat, and instead sweeping its neck around to gather food. For a while, another sauropod in Africa was considered to be a species of Barosaurus; today, however, it seems to be very clearly in its own genus, Tornieria, and actually far removed from both Diplodocus and Barosaurus (while still being in this closely related family group). So, for now, Barosaurus is only known from North America. These dinosaurs were distinctive long and slender sauropods, as opposed to their long and bulky cousins, the Apatosaurines, that they lived alongside.
~ By Meig Dickson
Sources Under the Cut
Allen, Eric Randall (Summer 2012). "Analysis of North American goniopholidid crocodyliforms in a phylogenetic context.” University of Iowa Research Online.
Apesteguía, S. 2005. Evolution in the hyposphene-hypantrum complex within Sauropoda. In K. Carpenter and V. Tidwell (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington 248-267.
Arldt, T. 1909. Die Dinosaurier [The dinosaurs]. Naturwissenshaftliche Rundschau 24(21):261-263.
Averianov, A. O., and T. Martin (2015). "Ontogeny and taxonomy of Paurodon valens (Mammalia, Cladotheria) from the Upper Jurassic Morrison Formation of USA" (PDF). Proceedings of the Zoological Institute of the Russian Academy of Sciences. 319 (3): 326–340.
Berman, D. S., and J. S. McIntosh. 1978. Skull and relationships of the Upper Jurassic sauropod Apatosaurus (Reptilia, Saurischia). Bulletin of Carnegie Museum of Natural History 8:1-35.
Bilbey, S.A. (1998). "Cleveland-Lloyd Dinosaur Quarry - age, stratigraphy and depositional environments". In Carpenter, K.; Chure, D.; and Kirkland, J.I. (eds.) (eds.). The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. pp. 87–120.
Bonaparte, J. F. 1986. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In K. Padian (ed.), The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary. Cambridge University Press, Cambridge 247-258
Bonaparte, J. F. 1986. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétosauridés) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentina) [The dinosaurs (carnosaurs, allosaurids, sauropods, cetiosaurids) from the Middle Jurassic of Cerro Cóndor (Chubut, Argentina)]. Annales de Paléontologie (Vert.-Invert.) 72(3):325-386.
Britt, B. B., and B. G. Naylor. 1994. An embryonic Camarasaurus (Dinosauria, Sauropoda) from the Upper Jurassic Morrison Formation (Dry Mesa Quarry, Colorado). In K. Carpenter, K. F. Hirsch, and J. R. Horner (eds.), Dinosaur Eggs and Babies 256-264.
Butler, R.J., P.M. Galton, L.B. Porro, L.M. Chiappe, D.M. Henderson, and G.M. Erickson. 2009. Lower limits of ornithischian dinosaur body size inferred from a new Upper Jurassic heterodontosaurid from North America. Proceedings of the Royal Society B 10.1098/rspb.2009.1494.
Caldwell, M. W.; Nydam, R. L.; Palci, A.; Apesteguía, S. N. (2015). "The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution". Nature Communications. 6: 5996.
Calvo, J. O., and L. Salgado. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. GAIA 11:13-33.
Carpenter K & Galton PM (2001). "Othniel Charles Marsh and the Eight-Spiked Stegosaurus". In Carpenter, Kenneth (ed.). The Armored Dinosaurs. Indiana University Press. pp. 76–102.
Carpenter, Kenneth (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131–138.
Carpenter, K. and Wilson, Y. 2008. A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Annals of the Carnegie Museum 76:227-263.
Carpenter, Kenneth (2018). Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado. Geology of the Intermountain West. 5: 227–244.
Carrano and Sampson, 2008. The phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 6, 183-236.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution 1-698
Charig, A. J. 1980. A diplodocid sauropod from the Lower Cretaceous of England. In L. L. Jacobs (ed.), Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert. Museum of Northern Arizona Press, Flagstaff 231-244
Chure, 2001. The second record of the African theropod Elaphrosaurus (Dinosauria, Ceratosauria) from the Western Hemisphere. Neues Jahrbuch für Geologie und Paläontologie Monatshefte. 2001(9).
Chure, Daniel J. (2001). "On the type and referred material of Laelaps trihedrodon Cope 1877 (Dinosauria: Theropoda)". In Tanke, Darren; and Carpenter, Kenneth (eds.) (eds.). Mesozoic Vertebrate Life. Bloomington and Indianapolis: Indiana University Press. pp. 10–18.
Chure, D. J., R. Litwin, S. T. Hasiotis, E. Evanoff, and K. Carpenter. 2006. The fauna and flora of the Morrison Formation: 2006. In J. R. Foster, S. G. Lucas (eds.), Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36:233-249
Coombs, W. P., and R. E. Molnar. 1981. Sauropoda (Reptilia, Saurischia) from the Cretaceous of Queensland. Memoirs of the Queensland Museum 20(2):351-373
Curtice, B. D. 1995. A description of the anterior caudal vertebrae of Supersaurus vivianae. Journal of Vertebrate Paleontology 15(3, suppl.):25A
Dalman, S.G. (2014). "New data on small theropod dinosaurs from the Upper Jurassic Morrison Formation of Como Bluff, Wyoming, USA". Volumina Jurassica. 12 (2): 181–196.
Demko, Timothy M.; Parrish, Judith T. (1998). "Paleoclimatic setting of the Upper Jurassic Morrison Formation". In Carpenter, Ken; Chure, Daniel J.; Kirkland, James I. (eds.). The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology 22 (1-4): 283-296.
Dodson, P. 1997. American dinosaurs. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 10-13
Ekart, Douglas D.; Cerling, Thure E.; Montanez, Isabel P.; Tabor, Neil J. (1999). "A 400 million year carbon isotope record of pedogenic carbonate; implications for paleoatmospheric carbon dioxide" (PDF). American Journal of Science. 299 (10): 805–827.
Engelmann, George F.; Chure, Daniel J.; Fiorillo, Anthony R. (2004). "The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation". In Turner, Christine E.; Peterson, Fred; Dunagan, Stan P. (eds.). Reconstruction of the Extinct Ecosystem of the Upper Jurassic Morrison Formation. Sedimentary Geology. Sedimentary Geology 167 (3-4): 297-308. 167. pp. 297–308.
Foster, John R. (1996). "Sauropod dinosaurs of the Morrison Formation (Upper Jurassic), Black Hills, South Dakota and Wyoming". Contributions to Geology, University of Wyoming. 31 (1): 1–25.
Foster, J.R. 2003. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. Bulletin 23.
Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327-329.
Foster, J. 2007. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 389pp.
Foster, J.R. 2009. Preliminary body mass estimates for mammalian genera of the Morrison Formation (Upper Jurassic, North America). PaleoBios 28(3):114-122.
Foster, J. (2018). "A new atoposaurid crocodylomorph from the Morrison Formation (Upper Jurassic) of Wyoming, USA". Geology of the Intermountain West. 5: 287–295.
Fraas, Eberhard (1908). "Ostafrikanische Dinosaurier". Palaeontographica. 55: 105–144.
Galianom, H., and R. Albersdörfer. 2010. A New Basal Diplodocoid Species, Amphicoelias brontodiplodocus from the Morrison Formation, Big Horn Basin, Wyoming, with Taxonomic Reevaluation of Diplodocus, Apatosaurus, Barosaurus and Other Genera. Dinosauria International (Ten Sleep, WY) Report for September 2010 1-41
Gallina, P. A., and S. Apesteguía. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentino de Ciencias Naturales, nuevo serie 7(2):153-166
Galton, P. M. 1977. The Upper Jurassic dinosaur Dryosaurus and a Laurasia-Gondwana connection in the Upper Jurassic. Nature 268(5617):230-232
Galton, P.M. & Powell, H.P. (1980). "The ornithischian dinosaur Camptosaurus prestwichii from the Upper Jurassic of England". Palaeontology. 23: 411–443.
Galton, P.M. (1981). Dryosaurus, a hypsilophodontid dinosaur from the Upper Jurassic of North America and Africa. Postcranial skeleton. Palaeontol. Z. 55(3/4), 271-312
Galton, 1982. Elaphrosaurus, an ornithomimid dinosaur from the Upper Jurassic of North America and Africa. Paläontologische Zeitschrift. 56, 265-275.
Galton PM, Upchurch P (2004). "Stegosauria". In Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition). University of California Press. p. 361.
Galton, P.M. (2010). "Species of plated dinosaur Stegosaurus (Morrison Formation, Late Jurassic) of western USA: new type species designation needed". Swiss Journal of Geosciences 103 (2): 187–198.
Galton, Peter M. & Carpenter, Kenneth, 2016, "The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen.", Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 279(2): 185-208.
Gillette, D.D. (1991). "Seismosaurus halli, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA". Journal of Vertebrate Paleontology. 11 (4): 417–433.
Gilmore, C.W. (1909). "Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and descriptions of two new species". Proceedings of the United States National Museum. 36 (1666): 197–332.
Harris, J. D., and P. Dodson. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 49(2):197-210
Harris, J. D. 2006. The axial skeleton of the dinosaur Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontology 49(5):1091-1121.
Hartman, S.; Mickey Mortimer; William R. Wahl; Dean R. Lomax; Jessica Lippincott; David M. Lovelace (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247.
Haughton, S. H. 1928. On some reptilian remains from the Dinosaur Beds of Nyasaland. Transactions of the Royal Society of South Africa 16:67-75
Hay, O. P. 1902. Bibliography and Catalogue of the Fossil Vertebrata of North America. Bulletin of the United States Geological Survey 179:1-868
Hendrickx, C, Mateus O. 2014. Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods, 03. PLoS ONE. 9:e88905., Number 3.
Holtz, Thomas R., Jr.; Molnar, Ralph E.; Currie, Philip J. (2004). Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 71–110.
Huene, F. v. 1908. Die Dinosaurier der Europäischen Triasformation mit berücksichtigung der Ausseuropäischen vorkommnisse [The dinosaurs of the European Triassic formations with consideration of occurrences outside Europe]. Geologische und Palaeontologische Abhandlungen Suppl. 1(1):1-419
Huene, F. v. 1909. Skizze zu einer Systematik und Stammesgeschichte der Dinosaurier [Sketch of the systematics and origins of the dinosaurs]. Centralblatt für Mineralogie, Geologie und Paläontologie 1909:12-22
Huene, F. v. 1927. Short review of the present knowledge of the Sauropoda. Memoirs of the Queensland Museum 9(1):121-126
Huene, F. v. 1927. Sichtung der Grundlagen der jetzigen Kenntnis der Sauropoden [Sorting through the basis of the current knowledge of sauropods]. Eclogae Geologica Helveticae 20:444-470
Huene, F. v. 1929. Los sauriquios y ornitisquios del Cretáceo argentino. Anales del Museo de La Plata, serie 2 3:1-196
Ibiricu, L. M., G. A. Casal, M. C. Lamanna, R. D. Martínez, J. D. Harris and K. J. Lacovara. 2012. The southernmost records of Rebbachisauridae (Sauropoda: Diplodocoidea), from early Late Cretaceous deposits in central Patagonia. Cretaceous Research 34:220-232
Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten [Overview of the vertebrate fauna of the Tendaguru beds]. Archiv für Biontologie 3:81-110
Janensch, Werner (1922). "Das Handskelett von Gigantosaurus robustus und Brachiosaurus brancai aus den Tendaguru-Schichten Deutsch-Ostafrikas". Centralblatt für Mineralogie, Geologie und Paläontologie. 1922: 464–480.
Jenkins, J.T. and J.L. Jenkins. 1993. Colorado's Dinosaurs. Denver, Colorado: Colorado Geologic Survey. Special Publication 35.
Joleaud, L. 1922. Les reptiles fossiles [Fossil reptiles]. Association Française pour l'Avancement des Sciences. Conférences. Compte Rendu de la 45e Session 49-66
Kenneth Carpenter; Peter M. Galton (2018). "A photo documentation of bipedal ornithischian dinosaurs from the Upper Jurassic Morrison Formation, USA". Geology of the Intermountain West. 5: 167–207.
Kirkland, J. I. 1997. Cedar Mountain Formation. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 98-99.
Kowallis, Bart J.; Christiansen, Eric H.; Deino, Alan L.; Peterson, Fred; Turner, Christine E.; Kunk, Michael J.; Obradovich, John D. (1998). "The age of the Morrison Formation" (PDF). In Carpenter, Ken; Chure, Daniel J.; Kirkland, James I. (eds.). The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology 22 (1-4): 235-260.
Ksepka, D. T., and M. A. Norell. 2010. The Illusory Evidence for Asian Brachiosauridae: New Material of Erketu ellisoni and a Phylogenetic Reappraisal of Basal Titanosauriformes. American Museum Novitates 3700:1-27.
Lockley, M.; Harris, J.D.; and Mitchell, L. 2008. "A global overview of pterosaur ichnology: tracksite distribution in space and time." Zitteliana. B28. p. 187-198.
Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional, Rio de Janeiro. 65 (4): 527–544.
Lucas S, Herne M, Heckert A, Hunt A, and Sullivan R. Reappraisal of Seismosaurus, A Late Jurassic Sauropod Dinosaur from New Mexico. The Geological Society of America, 2004 Denver Annual Meeting (7–10 November 2004).
Lucas, Spencer G.; Spielmann, Justin A.; Rinehart, Larry A.; Heckert, Andrew B; Herne, Matthew C.; Hunt, Adrian P.; Foster, John R.; Sullivan, Robert M. (2006). "Taxonomic status of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 149-161.
Lull, R. S. 1919. The sauropod dinosaur Barosaurus Marsh: redescription of the type specimens in the Peabody Museum, Yale University. Memoirs of the Connecticut Academy of Arts and Sciences 6:1-42.
Lull, R. S. 1924. Dinosaurian climatic response. In M. R. Thorpe (ed.), Organic Adaptation to Environment 225-279.
Lydam, R. L., Daniel J. Chure and Susan E. Evans (2013). "Schillerosaurus gen. nov., a replacement name for the lizard genus Schilleria Evans and Chure, 1999 a junior homonym of Schilleria Dahl, 1907" (PDF). Zootaxa. 3734 (1): 99–100.
Madsen, J. H., and W. E. Miller. 1979. the fossil vertebrates of Utah, an annotated bibliography. Brigham Young University Geology Studies 26(4):iii-147
Maidment, Susannah C.R.; Norman, David B.; Barrett, Paul M.; Upchurch, Paul (2008). "Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia)". Journal of Systematic Palaeontology. 6 (4): 367–407.
Mannion, P. D., P. Upchurch, O. Mateus, R. N. Barnes, and M. E. H. Jones. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology 10(3):521-551
Marsh, O. C. 1890. Description of new dinosaurian reptiles. The American Journal of Science, series 3 39:81-86.
Marsh, O. C. 1895. On the affinities and classification of the dinosaurian reptiles. American Journal of Science 50(300):483-498.
Marsh, O. C. 1896. The dinosaurs of North America. United States Geological Survey, 16th Annual Report, 1894-95 55:133-244.
Marsh, O. C. 1898. On the families of sauropodous Dinosauria. Geological Magazine, decade 4 3:157-158.
Marsh, Othniel C. (1899). "Footprints of Jurassic dinosaurs". American Journal of Science. 4 (7): 227–232.
Martin, John. 1987. Mobility and feeding of Cetiosaurus (Saurischia, Sauropoda) – why the long neck? pp. 154–159 in P. J. Currie and E. H. Koster (eds), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Boxtree Books, Drumheller (Alberta). 239 pages.
Mateus, O., & Antunes M. T. 2000. Ceratosaurus sp. (Dinosauria: Theropoda) in the Late Jurassic of Portugal. Abstract volume of the 31st International Geological Congress, Rio de Janeiro, Brazil.
Mateus, O. 2006. Late Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): a comparison. In J. R. Foster, S. G. Lucas (eds.), Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36:223-231.
Mateus, O. (2007). Notes and review of the ornithischian dinosaurs of Portugal. Journal of Vertebrate Paleontology. 27, 114A-114A., Jan: Society of Vertebrate Paleontology.
Mateus, O., & Tschopp E. (2013). Cathetosaurus as a valid sauropod genus and comparisons with Camarasaurus. Journal of Vertebrate Paleontology, Program and Abstracts, 2013. 173.
McIntosh, J. S. 1981. Annotated catalogue of the dinosaurs (Reptilia, Archosauria) in the collections of the Carnegie Museum of Natural History. Bulletin of Carnegie Museum of Natural History 18:1-67.
McIntosh, J. S. 1990. Sauropoda. In D. B. Weishampel, H. Osmólska, and P. Dodson (eds.), The Dinosauria. University of California Press, Berkeley 345-401.
McIntosh, J. S. 1997. Sauropoda. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 654-658.
McIntosh, J. S. 2005. The genus Barosaurus Marsh (Sauropoda, Diplodocidae). In K. Carpenter and V. Tidwell (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs. Indiana University Press, Bloomington 38-77.
Melstrom, K. M., M. D. D'Emic, D. J. Chure and J. A. Wilson. 2016. A juvenile sauropod dinosaur from the Late Jurassic of Utah, U.S.A., presents further evidence of an avian style air-sac system. Journal of Vertebrate Paleontology 36(4):e1111898:1-23.
Mezga, A., B. C. Tesovic, and Z. Bajraktarevic. 2007. First record of dinosaurs in the Late Jurassic of the Adriatic-Dinaridic carbonate platform (Croatia). Palaios 22(2):188-199.
Miller, W. E., J. L. Baer, K. L. Stadtman and B. B. Britt. 1991. The Dry Mesa Dinosaur Quarry, Mesa County, Colorado. In W. R. Averett (ed.), Guidebook for Dinosaur Quarries and Tracksites Tour, Western Colorado and Eastern Utah 31-46.
Mocho, P., R. Royo-Torres, and F. Ortega. 2014. Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society 170:875-916.
Moore, George T.; Hayashida, Darryl N.; Ross, Charles A.; Jacobson, Stephen R. (1992). "Paleoclimate of the Kimmeridgian/Tithonian (Late Jurassic) world: I. Results using a general circulation model". Palaeogeography, Palaeoclimatology, Palaeoecology. 93 (3–4): 113–150.
Nopcsa, B. F. 1928. The genera of reptiles. Palaeobiologica 1:163-188.
Olshevsky, G. 1992. A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia. Mesozoic Meanderings 2:1-268.
Parrish, J.T.; Peterson, F.; Turner, C.E. (2004). "Jurassic "savannah"-plant taphonomy and climate of the Morrison Formation (Upper Jurassic, Western USA)". Sedimentary Geology. 167 (3–4): 137–162.
Pickering, 1995a. Jurassic Park: Unauthorized Jewish Fractals in Philopatry. A Fractal Scaling in Dinosaurology Project, 2nd revised printing. Capitola, California. 478 pp.
Pritchard, A. C.; Turner, A. H.; Allen, E. R.; Norell, M. A. (2013). "Osteology of a North American Goniopholidid (Eutretauranosuchus delfsi) and Palate Evolution in Neosuchia". American Museum Novitates 3783 (3783).
Rauhut, O. W. M., K. Remes, R. Fechner, G. Cladera, and P. Puerta. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435:670-672.
Remes, K. 2006. A revision of the Tendaguru sauropod dinosaur Tornieria africana (Fraas) and its relevance for sauropod paleobiogeography. Journal of Vertebrate Paleontology 26(3):651-669.
Remes, K. 2007. A second Gondwanan diplodocid dinosaur from the Upper Jurassic Tendaguru Beds of Tanzania, East Africa. Palaeontology 50(3):653-667.
Romer, A. S. 1956. Osteology of the Reptiles, University of Chicago Press 1-77.
Romer, A. S. 1966. Vertebrate Paleontology, 3rd edition 1-468.
Royo-Torres, R., A. Cobos, A. Aberasturi, E. Espílez, I. Fierro, A. González, L. Luque, L. Mampel, and L. Alcalá. 2007. Riodeva sites (Teruel, Spain) shedding light to European sauropod phylogeny. Geogaceta 41:183-186.
Ruiz-Omeñaca, J. I., L. Piñuela, and J. C. García-Ramos. 2008. Primera evidencia de dinosaurios diplodocinos (Sauropoda: Diplodocidae) en el Jurásico Superior de Asturias (Noreña) [First evidence of diplodocine dinosaurs (Sauropoda: Diplodocidae) in the Upper Jurassic of Asturias (Noreña)]. In J I Ruiz-Omeñaca, L Piñuela and J C García-Ramos (eds), XXIV Jornadas de la Sociedad Española de Paleontología, 15-18 October 2008, Museo del Jurásico de Asturias (MUJA), Colunga, Spain, Libro de Resúmenes 191-192.
Russell, D., P. Beland, and J. McIntosh. 1980. Paleoecology of the dinosaurs of Tendaguru (Tanzania). Mem. Society Geol. France, N.S. 139:169-175.
Russell, D. A. 1984. A check list of the families and genera of North American dinosaurs. Syllogeus 53:1-35.
Saleiro, A., & Mateus O. (2017). Upper Jurassic bonebeds around Ten Sleep, Wyoming, USA: overview and stratigraphy. Abstract book of the XV Encuentro de Jóvenes Investigadores en Paleontología/XV Encontro de Jovens Investigadores em Paleontologia, Lisboa, 428 pp.. 357-361.
Salgado, L. 1993. Comments on Chubutisaurus insignis del Corro (Saurischia, Sauropoda). Ameghiniana 30(3):265-270.
Salgado, L. 1999. The macroevolution of the Diplodocimorpha (Dinosauria; Sauropoda): a developmental model. Ameghiniana 36(2):203-216.
Salgado, L., I. d. S. Carvalho, and A. C. Garrido. 2006. Zapalasaurus bonapartei, un nuevo saurópodo de La Formación La Amarga (Cretacico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina [Zapalasaurus bonapartei, a new sauropod from the La Amarga Formation (Lower Cretaceous), northwestern Patagonia, Neuquén province, Argentina]. Géobios 39:695-707.
Schultz, J. A; Bhart-Anjan S. Bhullar; Zhe-Xi Luo (2018). "Re-examination of the Jurassic mammaliaform Docodon victor by computed tomography and occlusal functional analysis". Journal of Mammalian Evolution. in press. doi:10.1007/s10914-017-9418-5.
Seebacher, Frank. (2001). "A new method to calculate allometric length-mass relationships of dinosaurs". Journal of Vertebrate Paleontology. 21 (1): 51–60.
Seeley, Harry G. (1869). Index to the fossil remains of Aves, Ornithosauria and Reptilia, from the Secondary system of strata arranged in the Woodwardian Museum of the University of Cambridge. Cambridge: Deighton, Bell and Co. pp. 143pp.
Sereno, P. C. 1997. The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences 25:435-489.
Smith, David K. (1998). "A morphometric analysis of Allosaurus". Journal of Vertebrate Paleontology. 18 (1): 126–142.
Smith, D. M., M. A. Gorman, J. D. Pardo and B. J. Small. 2011. First fossil Orthoptera from the Jurassic of North America. Journal of Paleontology 85(1):102-105.
Steel, R. 1970. Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology. Gustav Fischer Verlag, Stuttgart 1-87.
Sternfeld, R. 1911. Zur Nomenklatur der Gattung Gigantosaurus Fraas [On the nomenclature of the genus Gigantosaurus Fraas]. Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 8:398.
Suteethorn, S., J. Le Loeuff, E. Buffetaut and V. Suteethorn. 2010. Description of topotypes of Phuwiangosaurus sirindhornae, a sauropod from the Sao Khua Formation (Early Cretaceous) of Thailand, and their phylogenetic implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 256(1):109-121.
Swinton, W. E. 1970. The Dinosaurs, Wiley-Interscience, New York 1-331.
Tatarinov, L. P. 1964. Nadotryad Dinosauria. Dinozavry [Superorder Dinosauria. Dinosaurs]. In Y. A. Orlov (ed.), Osnovy Paleontologii [Fundamentals of Paleontology] 12:523-589.
Taylor, Michael P; Wedel, Mathew J (2013). "The neck of Barosaurus was not only longer but also wider than those of Diplodocus and other diplodocines". PeerJ. 1: e67v1.
Tidwell, V., Carpenter, K., and Miles, C., 2005, A reexamination of Morosaurus agilis (Sauropoda) from Garden Park, Colorado: Journal of Vertebrate Paleontology 25 (Supplement to 3):122A.
Tornier, G. 1913. Reptilia. Paläontologie [Reptilia. Paleontology]. Handwörterbuch der Naturwissenschaften 8:337-376.
Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. (2006). "Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations". Geological Society of America Abstracts with Programs. 38 (6): 7.
Tschopp, E., and O. Mateus. 2013. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. Journal of Systematic Palaeontology 11(7):853-88.
Tschopp, E., O. Mateus, and R. B. J. Benson. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3:e857.
Turner, Christine E.; Peterson, Fred (2004). "Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem—a synthesis". In Turner, Christine E.; Peterson, Fred; Dunagan, Stan P. (eds.). Reconstruction of the Extinct Ecosystem of the Upper Jurassic Morrison Formation. Sedimentary Geology. Sedimentary Geology 167 (3-4): 309-355.
Upchurch, P. 1995. The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London B 349:365-390.
Upchurch, P. 1995. Sauropod phylogeny and palaeoecology. In M. G. Lockley, V. F. dos Santos, C. A. Meyer, & A. P. Hunt (eds.), Aspects of Sauropod Paleobiology. GAIA 10:249-260.
Upchurch, P., P. M. Barrett, and P. Dodson. 2004. Sauropoda. In D. B. Weishampel, H. Osmolska, and P. Dodson (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 259-322.
von Zittel, K. A. v. 1890. Handbuch der Palaeontologie. I. Abteilung Paleozoologie. III. Band. Vertebrata (Pisces, Amphibia, Reptilia, Aves) [Handbook of Paleontology. Division I. Paleozoology. Volume III. Vertebrata (Pisces, Amphibia, Reptilia, Aves)] xii-900.
von Zittel, K. A. 1911. Grundzüge der Paläontologie (Paläozoologie). II. Abteilung. Vertebrata [Fundamentals of Paleontology (Paleozoology). Section II. Vertebrata]. Druck und Verlag von R. Oldenbourg, München 1-598.
Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. 861 pp.
Whitlock, J. A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161:872-915.
Wieland, G. R. 1920. The longneck sauropod Barosaurus. Science, New Series 51(1326):528-530.
Wild, Rupert (1991). "Janenschia n. g. robusta (E. Fraas 1908) pro Tornieria robusta (E. Fraas 1908) (Reptilia, Saurischia, Sauropodomorpha)". Stuttgarter Beiträge zur Naturkunde, Serie B: Geologie und Paläontologie. 173: 1–4.
Wilson, J. A., and M. B. Smith. 1996. New remains of Amphicoelias Cope (Dinosauria: Sauropoda) from the Upper Jurassic of Montana and diplodocoid phylogeny. Journal of Vertebrate Paleontology 16(3, suppl.):73A.
Wilson, J. A., and P. C. Sereno. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5. Journal of Vertebrate Paleontology 18(2 (suppl.)):1-68.
Wilson, J. A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136:217-276.
Xing, L., T. Miyashita, J. Zhang, D. Li, Y. Te, T. Sekiya, F. Wang and P. J. Currie. 2015. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. Journal of Vertebrate Paleontology 35(1):e889701:1-17.
#Barosaurus lentus#Barosaurus#Dinosaur#Diplodocid#Diplodocoid#Factfile#Palaeoblr#Jurassic#Herbivore#North America#Mesozoic Monday#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#Sauropod
332 notes
·
View notes
Note
I don't suppose we could known what highly obscure mammals' wiki pages this guy tried to vandalize? Just to browse their edit histories for a quick laugh. (I wouldn't bother asking anonymously normally, but I'd rather not deal with this guy's drama in my own inbox.)
Oh, he’s all over volaticotheres, meridolestidans, eutriconodonts, the like. He’s also all over dromaeosaur and troodont pages claiming they nested like megapodes (an idea that’s explicitly contradicted by the sources he cites; I and others have told him this multiple times, to his response of sticking his fingers in his ears).
I’ll also point to these wikipedia discussions, which are worth a good laugh.
76 notes
·
View notes
Note
If eutriconodonts can't fly why did they reach the Arctic before other true mammals did?
For one thing, we don’t know that they reached the Arctic before other mammals did. Haven’t you already had this conversation?

32 notes
·
View notes
Note
Addressed the points in regards to Ichthyoconodon
I see. Admittedly the majority of my proposed counters are fairly unlikely, but the aquatic and rafting ones still hold up.
We don’t know how different Ichthyoconodon’s body was from the confirmed glider clade but there’s at least a 30 million year ghost lineage from that clade. I don’t see why Ichthyoconodon’s teeth would necessarily have to be specialized, especially if it attained an amphibious lifestyle fairly recently. Again, river otter teeth look a lot like the teeth of most other mustelids, which had a similar diet to eutriconodonts, including volaticotheres. If it ain’t broke don’t fix it. As for why it’s so rare? This isn’t a perfect analogy but let’s look at the Kem Kem Formation, which has presumably been more heavily excavated and preserves a lot more fossils. We find a lot of isolated teeth. Lots of teeth from toothy pterosaurs, and only one non-tooth fossil. And this is a formation that overall has a lot more tetrapod fossils than the Anoual Syncline does. Maybe the Anoual Syncline just sucks at preserving mammals. And we can’t assume that the two isolated teeth came from the same individual, so we’ve got at least as many, if not more, Ichthyoconodon fossils known as any other mammals from that formation.
On rafting: of course we don’t have similar evidence for e.g. monkeys rafting across the Atlantic - the ocean that they would have fallen into is still ocean. Good luck finding fossils at the bottom of the Atlantic. And there are fossils of terrestrial animals - particularly compsognathids - that ended up dying at sea that don’t look particularly worn. You can still see all the little denticles on the teeth, which appear to retain their original texture. Whatever happened to them - including possibly falling off a raft of vegetation - could potentially happen to Ichthyoconodon.
So of all the counterarguments, the aquatic Ichthyoconodon argument holds up the most. I still don’t see any particularly compelling reason why flight is the most likely option for Ichthyoconodon other than phylogenetic proximity, and we all know how picky the fossil record is. We can’t really say much from two teeth but from what we can tell I’d personally still go with semiaquatic Ichthyoconodon.
And note these arguments also apply to Dyskritodon, whatever that was. Basically everything that applies to Ichthyoconodon could also apply to it. If you ask me, I’d say both were semiaquatic. If mink and river otters, or numerous pinnipeds, can coexist in the same place, I don’t see why they couldn’t.
10 notes
·
View notes
Text
Indotriconodon flies,baby!

By omegafreelancer
So learning that Indotriconodon magnus – an already impressive animal in being a large sized Mesozoic mammal and an unambiguous eutriconodont from the Maastrichtian – is also a volaticothere I naturally had to commission it in its likely volant form.
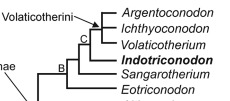
Volaticotheria phylogenetic tree by Bajpai et al 2024
Its debuting study also recover Sangarotherium and Eotriconodon as volaticotheres. The former in particular is the only mammal in an Arctic environment dominated by other synapsids (Averianov et al 2020), so it clearly must have been able to fly and migrate when other crown mammals couldn’t.
Things just keep looking up for volaticothere flight, which I already extensively argued for with the marine deposition and lack of dental wear of Ichthyoconodon, the early cosmopolitan distribution of the group, the fact that Volaticotherium‘s hand is largely missing and the fact that gliding mammals are primarily herbivorous (Luo 2017).
Naturally, I also used this image to forward my speculative volaticotheres over at Lemuria.
What a wonderous day.
References
Bajpai, Sunil; Rautela, Abhay; Yadav, Ravi; Wilson Mantilla, Gregory P. (2024-02-29). “The first eutriconodontan mammal from the Cretaceous (Maastrichtian) of India”. Journal of Vertebrate Paleontology. doi:10.1080/02724634.2024.2312234. ISSN 0272-4634.
Averianov, Alexander; Martin, Thomas; Lopatin, Alexey; Skutschas, Pavel; Schellhorn, Rico; Kolosov, Petr; Vitenko, Dmitry (2018-07-25). “A high-latitude fauna of mid-Mesozoic mammals from Yakutia, Russia”. PLOS ONE. 13 (7): e0199983. Bibcode:2018PLoSO..1399983A. doi:10.1371/journal.pone.0199983. ISSN 1932-6203. PMC 6059412. PMID 30044817.
Luo ZX, Meng QJ, Grossnickle DM, Liu D, Neander AI, Zhang YG, Ji Q (August 2017). “New evidence for mammaliaform ear evolution and feeding adaptation in a Jurassic ecosystem”. Nature. 548 (7667): 326–329. doi:10.1038/nature23483. PMID 28792934. S2CID 4463476.
#indotriconodon#eutriconodont#eutriconodonta#paleontology#palaeontology#paleoart#palaeoart#paleoblr#palaeoblr#mammal#cretaceous#maastrichtian
3 notes
·
View notes
Note
What do you think about Mesozoic mammals like eutriconodonts and multies?
I’m not an expert on them, but I do think that creatures like Repenomamus, Didelphodon and Purgatorius are cool, as is the fact that we had gliding mammals even back then!
2 notes
·
View notes
Photo

A Spiky Mammal from the Heyday of the Dinosaurs
This little beast speaks to mammalian success under the feet of the saurians
The classic view of the meek mammal eking out a living in a world dominated by dinosaurs is too simplistic, though. Mesozoic mammals are undergoing a renaissance – much like their saurian oppressors got in the 1970s – and have turned out to be far more varied than anyone expected.
Mammals of the Triassic, Jurassic, and Cretaceous were small, to be sure, but they counted swimmers, climbers, diggers, gliders, and more in their fuzzy family. They didn’t just live alongside the dinosaurs. They thrived. And among the latest beasties to be added to their ranks is a spiky mammal uncovered from the 125 million year old rock of Spain.
Named Spinolestes xenarthrosus by paleontologist Thomas Martin and colleagues, the mammal belonged to an extinct group called eutriconodonts...
(read more: Scientific American)
illustration by Oscar Sanisidro

28 notes
·
View notes
Photo

It’s finally time for “Almost-Living” Fossils Month!
There are various modern species that we call “living fossils” because they’ve survived for very long periods of geological time, but there are also plenty of long-lived and successful groups that didn’t quite manage to make it to the present day.
For all of August I’ll be doing six posts on this topic a week (taking a break on Sundays), and the basic criteria for entries are that a featured organism has to be part of a group that:
originated in the Mesozoic or earlier
existed for at least 50 million years
and survived through the end-Cretaceous mass extinction, only to go extinct later in the Cenozoic
So, let’s begin with...
Almost-Living Fossils Month #01 -- The Saint Bathans Mammal
Dating to the mid-Miocene, about 19-15 million years ago, this little mammal was discovered in the Saint Bathans fossil deposits in the South Island of New Zealand, and was probably the size of a large shrew, around 10cm long (4″). It doesn’t have an official scientific name yet, but its fragmentary remains might represent something very special.
Not only was it a terrestrial mammal living at a time when none were thought to inhabit New Zealand at all, but it also doesn’t seem to fit into any known group of mammals. It’s definitely not a placental or marsupial or monotreme, and it doesn’t match any of the other “archaic” groups -- so it could represent an entirely new lineage we didn’t even know existed.
At best it’s been classified as a theriiform, belonging somewhere between eutriconodonts and multituberculates in the mammal family tree. This means it would have to be part of a group with a very long ghost lineage going all the way back to the Late Triassic or Early Jurassic (~220-190 mya). Whatever it actually was, its ancestors were probably already present on New Zealand when the islands split away from Gondwana in the Late Cretaceous (~85 mya), and must have remained isolated there for tens of millions of years more.
It’s unknown how much closer to present day these mysterious archaic mammals actually survived, but it’s possible they were one of the casualties during the Middle Miocene disruption extinction about 14 million years ago, when the relatively warm climate suddenly cooled. And then land mammals were completely absent from New Zealand until the arrival of humans ~750 years ago.
(It’s also worth noting that the fossils of the Saint Bathans mammal were described over ten years ago, and our knowledge of ancient mammals and their evolutionary tree has changed a lot in that time. New studies or more fossil material might reveal the Saint Bathans mammal to actually be one of the very last survivors of an already-known group, but that would still make it an amazing and unique discovery.)
#almost living fossils month#science illustration#paleontology#paleoart#palaeoblr#saint bathans mammal#saint bathans fauna#theriiformes#incertae sedis#mammal#art#i'd especially like to see some comparisons done with all the mesozoic gliders we now know about#because the odd shape of its femur sounds potentially similar
255 notes
·
View notes
Last Seen Blogs

anayka-agarwal
Just Sharing

sweatybeautysoccerfreak
Unbetitelt

unabashedmagazinewizard
Untitled

rubski02
rubski

coveysongbirds
tigris snow's girlfriend