#diapsida
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
Tumblr Inc. has $15.1M in annual revenue.
Text
Monday Musings: Diapsida
One of the largest groups of vertebrates are the diapsids (two pairs of temporal fenestrae in the skull). Diapsids are nested in the larger clade sauropsida (reptiles, their ancestors and all their descendents).

The arrangement of the extra skull holes allows for bigger, stronger jaw muscles and enables the jaw to open wider.

A lesser ancestral characteristic is that of a long radius compared to the humerus.

Traditionally, diapsids were just modern reptiles and not their extinct relatives. In 1991, Laurin defined diapsida as "the most recent common ancestor of araeoscelidians, lepidosaurs, archosaurs and all their descendents. What does that mean to the non-scientist? Let's break it down.

Araeoscelidians were small lizard-like animals that lived in the Late Paleozoic Era. Often, this group is considered a sister group to diapsida but some do put them in the clade. They have slender legs, elongated tails and the two temporal fenestrae in their skulls.

Lepidosaurs are much better known by the lay person. This group includes all snakes, lizards and even the tuatara. Lepidosauria can be further split into two groups: squamata (lizards and snakes) and rhynchocephalia (tuatara is the only living member). Characteristics of lepidosauria include more flexible skulls (especially in squamates), the ability to self-amputate their tails (lost in some species_) and horny (keratinized) scales that make up the epidermis.
Other famous lepidosaurs include mosasaurs.

Archosaurs are also a fairly familiar group. Living members of this clade include crocodilians and birds.

The most famous members of this clade are probably the dinosaurs.

Archosaurs first appeared in the Triassic Period. Characteristics of the group include teeth set in deep sockets, mandibular fenestrae (hole in the mandible)

and a pronounced fourth trochanter (ridge on the femur where muscles attach),

Archosaurs move with an erect or semi-erect gait.

Recent phylogenetic studies place turtles somewhere between lepidosaurs and archosaurs with a common ancestor closer to archosaurs than lepidosaurs which makes them euryapsids, diapsids who secondarily lost their temporal fenestrae.
Now you know a little bit more about diapsids! Tune in tomorrow for some trivia!
#paleontology#fossils#dinosaur#fun facts#science#paleontologist#diapsida#lizards#snakes#tuatara#turtles#birds#crocodilians#science education#science terminology
22 notes
·
View notes
Text

Araeoscelis gracilis was a superficially lizard-like animal that lived during the mid-Permian, around 275 million years ago, in what is now Texas, USA. About 60cm long (~2'), it had a slender body, proportionally long legs, and a solidly-built skull with strong teeth, suggesting that it was a fast runner that specialized in cracking open the carapaces of thick-shelled prey.
It was one of the last known members of a lineage known as araeoscelidians, which are usually considered to be very early members of the diapsid reptiles – but a recent study has proposed they might have even more ancient roots than that, possibly being a branch of stem-amniotes instead.
———
NixIllustration.com | Tumblr | Twitter | Patreon
#science illustration#paleontology#paleoart#palaeoblr#araeoscelis#araeoscelidae#araeoscelidia#diapsida#reptile#amniote#art#lov the cronch
317 notes
·
View notes
Video
Munchique Wood-Wren by Jay Packer Via Flickr: Endemic to the western Andes of Colombia, this highly sought-after wood-wren is found at higher elevations than the similar Gray-breasted Wood-Wren. Montezuma Road, Tatamá National Park, Colombia
#Animalia#Amniota#Aves#Reptilia#Vertebrata#Archosauromorpha#Chordata#Diapsida#Tetrapoda#Munchique Wood-Wren#Colombia#Troglodytidae (Wrens)#Passeriformes#South America#Animals#Birds#Henicorhina negreti#flickr
12 notes
·
View notes
Text

Gracilicollum
Gracilicollum — рід ймовірних таністрофеїдних архозавроморфів з середньотріасової (анізій) формації Гуаньлін у Китаї. Рід містить один вид, G. latens, відомий за черепом і частковою шиєю.
Повний текст на сайті "Вимерлий світ":
https://extinctworld.in.ua/gracilicollum/
#gracilicollum#triassic#archosauromorpha#china#reptiles#ua#diapsida#asia#art#paleontology#paleoart#prehistoric#animals#animal art#extinct#article#digital art#fossils#illustration#палеоарт#палеонтологія#ukraine#ukrainian#україна#мова#арт#українська мова#наука#китай#тварини
36 notes
·
View notes
Note
Birds are class Aves.
Sure, under Linnaean taxonomy. But, well,
A) Linnaeus was a eugenecist so his scientific opinions are suspect and his morality is awful
B) he didn't know about evolution
C) he didn't know about prehistoric life
so his classification system? Sucks ass. It doesn't work anymore. It no longer reflects the diversity of life.
Instead, scientists - almost across the board, now - use Clades, or evolutionary relationships. No rankings, no hierarchies, just clades. It allows us to properly place prehistoric life, it removes our reliance on traits (which are almost always arbitrary) in classifying organisms, and allows us to communicate the history of life just by talking about their relationships.
So, for your own edification, here's the full classification of birds as we currently know it, from biggest to smallest:
Biota/Earth-Based Life
Archaeans
Proteoarchaeota
Asgardians (Eukaryomorphans)
Eukaryota (note: Proteobacteria were added to an asgardian Eukaryote to form mitochondria)
Amorphea
Obazoa
Opisthokonts
Holozoa
Filozoa
Choanozoa
Metazoa (Animals)
ParaHoxozoa (Hox genes show up)
Planulozoa
Bilateria (all bilateran animals)
Nephrozoa
Deuterostomia (Deuterostomes)
Chordata (Chordates)
Olfactores
Vertebrata (Vertebrates)
Gnathostomata (Jawed Vertebrates)
Eugnathostomata
Osteichthyes (Bony Vertebrates)
Sarcopterygii (Lobe-Finned Fish)
Rhipidistia
Tetrapodomorpha
Eotetrapodiformes
Elpistostegalia
Stegocephalia
Tetrapoda (Tetrapods)
Reptiliomorpha
Amniota (animals that lay amniotic eggs, or evolved from ones that did)
Sauropsida/Reptilia (reptiles sensu lato)
Eureptilia
Diapsida
Neodiapsida
Sauria (reptiles sensu stricto)
Archelosauria
Archosauromorpha
Crocopoda
Archosauriformes
Eucrocopoda
Crurotarsi
Archosauria
Avemetatarsalia (Bird-line Archosaurs, birds sensu lato)
Ornithodira (Appearance of feathers, warm bloodedness)
Dinosauromorpha
Dinosauriformes
Dracohors
Dinosauria (fully upright posture; All Dinosaurs)
Saurischia (bird like bones & lungs)
Eusaurischia
Theropoda (permanently bipedal group)
Neotheropoda
Averostra
Tetanurae
Orionides
Avetheropoda
Coelurosauria
Tyrannoraptora
Maniraptoromorpha
Neocoelurosauria
Maniraptoriformes (feathered wings on arms)
Maniraptora
Pennaraptora
Paraves (fully sized winges, probable flighted ancestor)
Avialae
Avebrevicauda
Pygostylia (bird tails)
Ornithothoraces
Euornithes (wing configuration like modern birds)
Ornithuromorpha
Ornithurae
Neornithes (modern birds, with fully modern bird beaks)
idk if this was a gotcha, trying to be helpful, or genuine confusion, but here you go.
all of this, ftr, is on wikipedia, and you could have looked it up yourself.
679 notes
·
View notes
Text
Just found this amazing paper reconstructing what kinds of animal sounds you would hear in prehistoric landscapes:
Stridulating insects, including crickets, performed the first terrestrial twilight choruses during the Triassic. The twilight chorus was joined by water boatmen in the Lower Jurassic, anurans [frogs] in the Upper Jurassic, geckoes and birds in the Lower Cretaceous, and cicadas and crocodilians in the Upper Cretaceous. Parallel evolution of defensive stridulation took place multiple times within Malacostraca, Arachnida and Coleoptera [possibly from the Devonian]. Parallel evolution of defensive and courtship-related sound production took place in Actinopterygii [bony fish], possibly as early as the Devonian. Defensive vocalisations by tetrapods probably did not appear until their predators acquired tympanic ears in the Permian. Tympanic ears appeared independently in Diadectomorpha, Seymouriamorpha, Parareptilia, Diapsida and derived Synapsida. Crocodilians and birds acquired vocal organs independently, and there is no anatomical evidence for vocal ability in bird-line archosaurs basal to the avian clade Ornithothoraces. Acoustic displays by non-avian dinosaurs were therefore probably non-vocal. Other aspects of the evolution of acoustic behaviour in these and other lineages are also discussed.
22 notes
·
View notes
Text

Cockatrice (Artwork by Yuujinner)
Kingdom: Animalia
Phylum: Chordata
Class/Clade: Reptilia (Sauropsida)
Clade: Diapsida
Clade: Archosauria
Clade: Dinosauria
Clade: Saurischia
Clade: Eusaurischia
Clade: Theropoda
Clade: Neotheropoda
Clade: Averostra
Clade: Tetanurae
Clade: Avetheropoda
Clade: Coelurosaria
Clade: Maniraptoriformes
Clade: Maniraptora
Clade: Pennaraptora
Clade: Oviraptorosauria
Clade: Edentoraptora
Superfamily: Caenagnathoidea
Family: Caenagnathidae?
Subfamily: incertae sedis
Genus: Ornithosaurus
Species: O. necrophilus (“death-loving bird lizard”)
Ancestral species: possibly Microvenator celer
Temporal range: late Pleistocene to recent (87,000 kya - present)
Information:
A ravenous scavenger, the cockatrice is by no means at the top of its food chain, though its uniquely offensive, musky odor, ear-splitting vocalizations, and proclivity for traveling in large groups called flocks make it a creature which few predators wish to tolerate. Add onto this its territorial aggression, and you have what may be Archaeonesia’s most detested scavenger. Cockatrices use their superb sense of smell to detect carrion from several tens of miles away, primarily feeding on the carcasses of various reptilian and mammalian megafauna, sometimes flocking around fresh kills made by larger predators and using their sheer number to overwhelm the carnivore into relinquishing its kill. Though it usually eats carrion, it is also classified as an opportunistic feeder, readily going after small vertebrates. Found primarily in the Arava Desert and the surrounding grasslands in the western half of the Isle of Perils, this medium-sized oviraptorosaur is known all throughout the Isle of Perils, including its central mountain range, making it one of the few non-avian dinosaurs to live in that region. It is also one of the few non-avian dinosaurs to actively seek out human settlements, particularly to feed on discarded scraps of food. Actively seeking out human settlements, it is known to scavenge from trash heaps and refuse bins, which make it a local pest in some areas. Entire flocks of these animals, as many as 40 individuals sometimes, may swarm landfills. Similarly, these creatures will use their sheer number of overwhelm larger carnivores into relinquishing kills before greedily tearing into their spoils. A pecking order can be observed amongst these animals, typically in which the largest male gets first pickings on the corpse. When feeding on carrion, as gruesome as it may be, they will typically eat away at the orifices first before hollowing out the cadaver. Due to its exceptionally strong stomach acids being able to kill most bacteria, it can eat carrion which most other scavengers would otherwise find too putrid or dangerous to consume. Attracted to shiny objects for the purposes of adorning their nests with them, they have been known to steal jewelry, though those which live farther from human settlements may instead use quartz and other naturally occurring crystals to adorn their nests. These animals are exceptional jumpers, being able to clear fences nearly 12 feet all and jump nearly 25 feet in a single bound. Exceptionally territorial in nature, groups may mark trees and rocks with a pair of scent glands behind their ears, which produce the foul musky odor typically associated with the animal. As these animals are quite social, their ability to recognize patterns (and more specifically faint color patterns and facial differences) allow them to differentiate between one another with remarkable ease. They can also recognize human faces with exceptional accuracy. Grooming behavior is well-documented, and like primates, it plays an important role in establishing social relations. Primarily diurnal, these animals rely on scent and eyesight to find food, and typically, a few individuals will venture away from the nesting grounds at a given time to locate food before they’ll go back and alert the others of its location, utilizing what is sometimes described as an elaborate“dance”, consisting of many different vocalizations, as well as head and body movements, to communicate location, much in the same way honeybees do. As the many environments it lives in are teeming with predators, a few individuals will take shifts throughout the night to watch the nesting grounds while the others sleep. A pouch at the base of the neck, commonly called a crop, allows the animal to store food before digesting it, though it serves a dual function of allowing it to transport food back to the nest to feed its offspring.
Though a given flock of cockatrices may not necessarily consist of entirely closely related individuals, it is more common than not for a flock to consist of a set of parents or grandparents and several generations of offspring. During the beginning of the dry season, around early December, the males’ colors will become substantially more flashy and eye-catching, his wattle flushing a bright maroon and violet color and the undersides of his wings flushing a pink hue, and although related species are known to engage in mock fights as part of mating displays, this species instead relies on a less violent method of winning approval from the females they wish to court: designing the most colorful display. A male will create a nest and adorn it with the most colorful materials he can found, anything from flowers and fruits to rocks and crystals. However, this is only part of the courtship ritual. While a bright nest may earn some initial interest from female suitors, it is what he does next that determines his success: performing an elaborate dance, sometimes with a shiny rock clutched in his beak, he will angle his head up towards the sky, revealing his brightly-colored wattle and wings. High-stepping in a circle around her, his throat will undulate to make a deep, rattling bellow, beating his wings and jumping up and down to keep her attention. If she accepts, she will join him in this dance and copulation begins. Cockatrices mate for life, and in 1.5 months time, she will lay a clutch of 2-4 blue eggs in the nest, and for the 5 weeks it will take for them to hatch, she will not leave the nest, the male fetching her food and water via his crop. When the young are born, they are, in a rare exception amongst non-avian theropods, altricial, being born nearly featherless and unable to walk for the first few weeks of life. By a month old, they will be able to walk. By a year, they will have reached half their adult size, being large enough to join their parents in the search for food. By 2 years, they will reach adult size, and at around 3.5 years, they will have gained their adult plumage and will reach sexual maturity. Many may choose to stay with their parents’ flock, though some may go off and form flocks with other young cockatrices. If they’re lucky, a cockatrice may expect to live 20-30 years.
Around the size of a cassowary, this species is around 5.6 feet tall, roughly 9-10 feet in length, and weighs around 200 lbs on the heavier side. There is no notable sexual dimorphism between species. The naked head is highly fluorescent, the neck being reddish yellow and the wattle/fleshy growths on its face being yellowish-orange and bluish-purple. The beak is red and the eyes are white. Plumage is white on the body and most of the wings, though near the base of the neck, the tail, and the wing feathers, the plumage starts to turn black, with the wing plumage having many beige spots along their length. Its legs are yellowish-gray.
Long-renowned for its dissonant calls, this species generally communicates with others of its kind with rasps, shrill humming, and a sound variously called “bleating” or “bugling”. Territorial calls consist of loud, deep booms which rumble across the land. However, it may hiss or honk if aggravated or in an attempt to intimidate and size up other scavengers/carnivores, and it has a characteristic shrieking whoop referred to by some as a “dinner bell call” to other cockatrices that food has been located.
Much in the same way that vultures are viewed as unclean and malevolent animals in Western society, so, too, is the cockatrice in Xenogaean society, made dually ironic for the fact that vultures also exist in the region, albeit typically in more montane environments. Long seen as a bad luck omen, stumbling across a dead cockatrice was said to signal impending disaster, particularly famine or drought, and in fact, it was said that if one did stumble across one, or managed to kill one, they were to immediately cremate it and spread its ashes in a river. Nonetheless, it does appear in some heraldic imagery and was venerated amongst some indigenous peoples in the region, particularly to the southeast. It was said the Bronze Age Aravan King, Kuntapurexa, infamous for his brutal conquests across the Isle of Perils, was followed by a horde of cockatrices which reaped the benefits of his conquests, feeding on the corpses of those he and his men killed as they went from village to village pillaging and marauding. The deafening sounds of these animals from afar was therefore used by some villagers as a way to determine how close Kuntapurexa and his men were to their settlement and therefore whether or not to abandon the town. How true this was, however, remains up to speculation, as no surviving historical records seem to confirm if this was a true account or not, with the possibility of it being a tall tale being rather likely. That said, if one can get past the animal’s revolting smell and dietary habits, a tame cockatrice makes for an exceptional companion animal, being exceptional at navigating, tracking, and retrieving items and trinkets, and in times past, some would use these animals to discretely transmit messages across long distances in a similar manner to messenger pigeons. On top of that, its affectionate nature towards those it’s acquainted with makes it decent as a pet as well, minus its food requirements. In fact, while some cities actively try to exterminate or otherwise relocate cockatrices within their walls, others may actively promote breeding programs for the animals in an effort to reduce waste in landfills. Despite being classified as a caenagnathid oviraptorosaurian, this placement is tentative: though its skull anatomy and genetic data would seem to support an inclusion amongst the Caenagnathidae or at least closer to the Caenagnathidae than the Oviraptoridae, the anatomy of its arms (and its wrists in particular) is exceptionally basal, more akin to that of therizinosaurs or ornithomimosaurs than to that of other oviraptorosaurs. Amongst an indigenous group in the Arava Desert region known as the Nge'echets, the cockatrice was seen as an embodiment of the desert itself, almost a god in its own right, far contrary to how their Xenogaean-speaking neighbors viewed the animal. As such, offerings were left out to the animals as a way of asking for safe passage from one oasis to the next as part of their migratory lifestyle. Nonetheless, amongst all native cultures in the region, the consumption of this animal’s meat is considered taboo due to its scavenging lifestyle. In lieu with its scavenging lifestyle, flocks of these animals may follow sick or injured animals for miles, waiting for them to collapse before finishing them off, hence it was long said that spotting a cockatrice behind oneself was a sign that death was on one’s doorstep. In some regions, they are also associated with the Xenogaean death goddess, Yerakiya, seen as either her messengers or even as a form she herself takes in the world of the living. Bones of this animal date back to the late Pleistocene, around 87,000 years ago, and fossil member of the genus are known as far back as the Miocene. A smaller closely-related species found on an offshore island, the basilisk (O. insularis), went extinct in the 18th century due to the introduction of pigs by British colonists. With around 2,000,000 mature adults in the wild, populations appear to be stable but declining in certain areas.
#scifi#fantasy#speculative evolution#novella#scififantasy#speculative biology#speculative fiction#speculative zoology#worldbuilding#creature art#oviraptorosauria#cockatrice#sci fi#scifi worldbuilding#fantasy worldbuilding#fantasy creature#scifi creature#dinosaurs#dinosaur#spec evo
13 notes
·
View notes
Note
Interesting !! Is there like a specific classification system you're going by for the poll order? I know you mentioned the Linnaean system somewhere, does that just mean you're sort of going by rank and ignoring phylogeny (for lack of a better wording)?
Also, lichen likers (almost wrote lichers) are on their own. Bestfungi and bestalgae can handle that 😭😭😭
Once I get to classes, I’m not sure. I think it will be mostly linnaean, but I may substitute some clades that make more sense (such as using Diapsida or Sauria instead of splitting birds from the other reptiles, or have a seperate poll for Lepidosaurs and Archosaurs). I’ll just be playing it by ear. 😅
7 notes
·
View notes
Text
Dinosaurs are not extinct.
The fact that Dinosaurs, the rulers of Mesozoic earth are extinct is not at all a 'news' to you, right?
Yeah, I mean considering you or your distant cousins belong to the Holocene earth and actively taking part in this Anthropocene, you have been technically taught right since your childhood, that YES! DINOS, THE GIANT LIZARDS ARE EXTINCT. And, even if I write something on this, won't make a big deal. But what will make quiet a big deal is, if I write and actually go onto claim that NO! DINOSAURS AREN'T EXTINCT! Now this is what that sixth grader would say, "Yes, I want more of this... go on I'm listening."
By now, you might be thinking that this is a big statement I have made. But what if I tell you, go and ask this same question to a paleontologist or maybe some evolutionary biologist. Trust me, try it out and you'll see, it's not much of a shock to them as it was to you. Dinosaurs are not extinct, they know it. But do they say it? NO (and this is what this blog is all about)
You see, in the world of Paleontology or (to be more precise)Dinosaur Paleobiology there are two very vaguely defined things. And by saying, 'very vague' it's actually quiet serious. I mean if at least one of these two would have been crystal clear to us, I wouldn't have been writing this blog today.
First, the very meaning of the word Dinosaur! yes... it literally means "A Giant Lizard" (as you all might already know). Why Sir Richard Owen, why did you do this? I mean there's a whole different story to this though, but yeah the great paleontologist of his time could have come up with something more better and clearer.
Now, if you are that average sixth grader wondering what's actually wrong with A GIANT LIZARD, well let me shatter your childhood learnings and say that, dinosaurs are related to lizards but not that related enough to actually call them lizards!
Avoiding, scientific stuffs here (you'll get bored otherwise) let me get this straight. Assuming you are aware of a phylogenetic tree, look at this below -
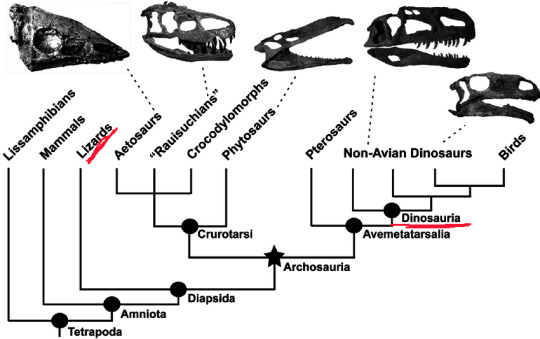
Just look at where Lizards and the group of Dinosauria or Non Avian Dinosaurs are! In our planet's great evolution story (with regards to Geological Time Scale of course), lizards and dinosaurs both went on to follow separate paths of development.
Now the sixth grader might argue, "Oh cmon! but they do have a common ancestry at some point right, like mentioned here as Diapsida?" Well its true, but going with this, it all comes down to that one single-celled microorganism or maybe that RNA molecule made of just proteins from which life eventually began and the fact then becomes, YOU are also a lizard or fish or any insect. Not so happy with that ain't you. Trust me, my brain's okay digesting this but you or an average sixth grader won't be proud calling themselves a disgusting house fly :)
This is what I call 'A Game of Perspective'.
Perspective, to actually define events with regards to our evolutionary history. This is... what I feel, the most difficult thing to do in paleontology, apart from the fact that you define something one day and tomorrow you dig something out related to your newly defined 'thing' which actually makes you rewrite the whole definition, previous day going in vain.
Hence, the giant lizard, based on the above phylogenetic tree, does live. It's not at all extinct. Komodo dragon, take this for an example if you aren't satisfied with your house lizard. Now, this obviously doesn't bring back the famous Tyrannosaurus Rex or Velociraptor right? They are indeed extinct. But, if you are attentive enough to actually see 'Birds' in the above phylogenetic tree branching out of the Dinosauria group, ask yourself, "Is the chicken, that I love to eat from KFC extinct?"
The answer is an undebated NO and this is where the 'second vague' thing comes in - Where to draw the line and define birds as a whole separate species?

Look at a closer division of Dinosauria in the above figure and you will see that 'Aves' has actually branched out from 'Theropoda', which itself has branched out from 'Saurischia'. Aves, is what you might already know, refer to the birds. Now the main point of confusion is at what point in evolutionary history did a chicken become a chicken from its theropod descendants. The game of perspective comes up here, and we look into the definition of birds with regards to theropods (group of Dinosauria), because we know that they are their evolutionary successors. (look at the above figures again if you still don't believe)
Birds as Archeopteryx and all of its descendants.
Pretty cool right? I mean if you follow paleontology to some extent you might know that Archeopteryx is the missing link fossil between a bird and a dinosaur. But is it a bird or a dinosaur?
More precise - Is Archeopteryx a bird or a theropod?
The issue here is, recent studies have shown that Archeopteryx is more related to Dromaeusaurids (a small clade of feathered dinosaurs) than modern day birds.
2. Birds as feathered dinosaurs.
As good as it may sound, feathers! It is the most confusing body part of a Dinosaur, because more and more fossils show that they had feathers attached to them. Even your favorite T-Rex is now believed to have it! (Note - Jurassic Park is just a movie, there's a whole lot more to dinosaurs than to just get chased by them in a park)
3. Birds as flying dinosaurs.
Again, it is difficult to determine exactly which dinosaurs were capable of flying (as opposed to simply gliding). Also from a modern perspective, penguins for instance don't fly. But they are birds!
4. Birds as crown dinosaurs, meaning the last common ancestor of all extant birds and its descendants.
This is somewhat the most favored definition by paleontologists till date, but the problem here is, it ignores many feathered and flying dinosaurs that are more closely related to modern birds than to Archaeopteryx as birds.
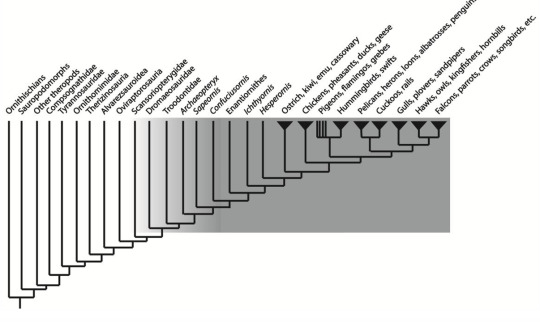
Just look at how the highlighted region fades as we go from modern birds towards Archeopteryx and so on.
So, YES... saying that the Ornithischian group of dinosaurs is extinct, isn't wrong at all. Even the Sauropod group of Saurischian dinosaurs are extinct. But the Theropods, the same evolutionary line of Tyrannosaurus Rex isn't extinct, unless paleontologists can actually draw the line to separate modern day birds from them.
Paleontologists are still trying to figure this line out and believe me, this is quite challenging. I would like to quote here, studying what's underneath is much more difficult than something just above your head. You see, your goal isn't far in paleontology... it's hidden and that's what makes it more difficult.
Now, is it even necessary to actually draw this line. I'm afraid it is, otherwise blogs like this will come up more and disturb the already disturbed lives of paleontologists, dinosaur paleobiologists out there.
And... till the time this line is drawn, respect the chickens that you eat as they are your living dinosaurs. (I mean, I am not promoting veganism here, but just respect them, yeah... can't think of anything else)
Figure References - Figure 1 (in order) taken from Macroevolutionary patterns in the evolutionary radiation of archosaurs (Tetrapoda: Diapsida) by Stephen L Brusatte | September 2010 Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101(3-4) Figure 2 (in order) taken from www.opengeology.org | Image by Callan Bentley Figure 3 (in order)taken from Dino101: Dinosaur Paleobiology 200/201 University of Alberta | Modified from Hackett et al.2008
If you have made it this far, thanks a lot. Feel free to ask me any dumb thing. Trust me, dumb questions often lead to great discoveries!
Anyways, this is it for my first blog.
For the love of Earth Science :D Byeee
23 notes
·
View notes
Text
Massive Winged Dragons

Diagram by David E. Alexander, showing various species of Permian-Triassic gliding reptiles. Compare these wings to the modest wings of modern Draco lizards.
Folks familiar with paleontology might have heard of the kuenheosaurids and weigeltisaurids, two lineages of extinct gliding reptiles superficially similar to modern Draco lizards. The former were apparently stem-lepidosaurs distantly related to modern squamates and tuataras, while the latter are a much older clade with no living relatives. Both groups have wings supported by rib processes, but whereas kuenheosaurids have typical rib-wings like their modern gliding lizard analogues those of weigeltisaurids are actually elements seperate from the ribs proper, allowing them to both have functional ribcages and rib wings!
Though the exact positioning of these elements is still unclear.
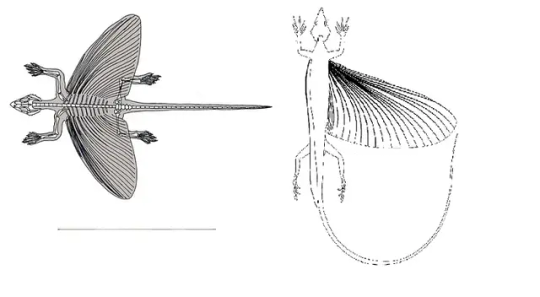
Two diagrams of Coelurosauravus, the first with the wing support elements aligned along the ribs and the latter radiating from the armpits.
As you can see, the wings of these animals tend to be proportionally very large for gliders, as well as proportionally elongated. Those of modern Draco lizards are much smaller and rounder, though those of the extinct lizard Xianlong are almost as large(Li 2007). Furtheremore, kuenheosaurids at least co-existed with another rib-glider, Mecistotrachelos, which had wing proportions more similar to those of modern Draco lizards in contrast with the proportionally massive wings for the former (Fraser 2007).

Mecistotrachelos by Cerri Thomas, displaying wing proportions similar to those of modern Draco lizards.
Unlike volaticotheres and non-avian theropods, I’m not going to argue that these were powered flyers, since even weigeltisaurids lack a sternum to which downtroke muscles would be attached (unless they had them attached to the back, like a reverse neornithine condition?).
However, I do think that this hints at far more aerial habits than modern Draco lizards, perhaps relying on thermals or other air currents to soar for long distances. Notably, several kuenheosaurids have different wing shapes (see diagram above), having radiated much as flying birds long after them.
The era of these large winged flyers also coincidentally ends with the arrival of pterosaurs to the scene, with only Xianglong approaching similar wing proportions. Perhaps these gliders occupied niches latter taken by true flying vertebrates, which to me suggests that they weren’t functionally analogous to modern gliding vertebrates.
Food for thought.
References
Pi-Peng Li; Ke-Qin Gao; Lian-Hai Hou & Xing Xu (2007). “A gliding lizard from the Early Cretaceous of China” (pdf). Proceedings of the National Academy of Sciences of the United States of America. 104 (13): 5507–5509. doi:10.1073/pnas.0609552104. PMC 1838464. PMID 17376871.
Fraser, N.C.; Olsen, P.E.; Dooley, A.C. Jr.; Ryan, T.R. (2007). “A new gliding tetrapod (Diapsida: ?Archosauromorpha) from the Upper Triassic (Carnian) of Virginia” (PDF). Journal of Vertebrate Paleontology. 27 (2): 261–265. doi:10.1671/0272–4634(2007)27[261:ANGTDA]2.0.CO;2. ISSN 0272–4634.
5 notes
·
View notes
Photo
That is... ...a lot of separation to consider those two things (dinosaurs and birds) the same. Remember that cladistic connection express two things: time and feature similarity. It doesn’t even necessarily mean the latter descended from the former, although it does mean the scientific community considers that line of descent to be the most plausible (assuming the particular chart being consulted is the most up-to-date, which the chart you quoted sounds like it is). Nature rarely draws lines without smudging them, so a sufficiently detailed cladistics chart (more properly known as a cladogram, a graph representing how clades - individual species or closely-connected groups of species - relate across time) should have transitional species (and larger groups of species, which then form clades of their own) between two large categories of animals that are significantly different in nature. There is also the question of what size groups “birds” and “dinosaurs” are. Would one say “birds” are like “primates”, “mammals” or something else? All make different presumptions about when something ceases to be a bird, and similar notions apply to dinosaurs too. The popular and scientific definitions of both may not 100% correspond (indeed, the “humans are fish” meme relies on it). Thus, the options for cladistic dinosaur-bird relationships, as I see them are: - There are clades between dinosaurs and birds that are dinosaurs in some respects and birds in others. This is the most likely option. - There are clades that are dinosaurs in some respects and a secret third thing in others, followed by clades that are a secret third thing in some respects and birds in others (note that these clades probably also have some inheritances from dinosaurs, for the same reason as humans have an aspiration risk inherited from fish without credibly being classifiable as fish or “land fish”) This exists for ancestors of dinosaurs and birds as I type (May 2023). The “Archosauromorpha” category exists for animals that are more like Archosauria than Lepidosauria and was created when the molecular record was shown no longer to support a direct descendency between Archosauria and their parent clade (Diapsida). This is still being researched, so if this becomes a World Heritage Post and cladograms show something else when you read this, that’s why.
- Something in the detailed internal classification of clades between dinosaurs and birds is slightly awry - not enough to change the basic descendent relationship, but enough that something important about how they connected is not yet fully understood and encoded into the system. This is the first option that requires any changes whatsoever to the currently-understood cladistic chart, and in this case requires rearranging a few of the less-evidenced lines to clarify which feature similarities changed when. Compatible with but not obliging of the first two options. - The features that distinguish a dinosaur or bird change in such a way that more of the intermediate cases are grouped into one or the other. Compatible with but not obliging of all options above. - Either the definition of “dinosaur” or “bird” is determined by the scientific community to be dominant over the other one. I’ve seen children’s dinosaur books in the 1990s do a version of this, and loudly insist that pterosaurs cannot possibly be considered dinosaurs purely because they flew, but should instead be their own category. This is the first one that does not suggest “birds are birds rather than members of a category of the same sort of level as dinosaurs” - “Dinosaur” is defined formally as an inheritable subset of “reptile” (which would make it impossible for any clade linked to it to be deemed anything other than a dinosaur) and loses its more common definition of being a category of a specific type of animal during the Mesozoic era. This is unlikely because fish already aren’t defined this way, and unless I’ve missed something neither are dinosaurs. This is, however, the option that would allow birds to be classified as dinosaurs. - Someone points out that as neither “dinosaur” nor “bird” directly appears on a properly-formed cladogram, thus cladograms should not be used to describe the relationship between them (only the relationship between clades that in turn may be described as part of the more commonly-used terms). This is unlikely to happen because cladistics intuitively would be expected to describe the relations between different types of animal. - (least likely) there was an error somewhere in this cladistic line large enough that birds don’t belong in the same cladistic line as dinosaurs. There is so much evidence of the ancestral link by this point that one might as well argue that humans are dinosaurs as suggest this option for dinosaurs and birds.

CLADISTICS ruined my life
114K notes
·
View notes
Text

Coelurosauravus (berarti "kakek kadal berongga") adalah sebuah genus reptil Diapsida basal, dengan struktur seperti sayap khusus yang memungkinkannya meluncur di udara. Ini adalah batang seperti struktur dengan kulit yang membentang di atasnya. Batang tulang ini bukan perpanjangan tulang rusuk, melainkan tulang dermal yang baru berevolusi.
0 notes
Text

Drepanosaurs were a weird little group of tree-climbing Triassic reptiles with prehensile claw-tipped tails, chameleon-like bodies, humped backs, grasping feet, long necks, and somewhat bird-like skulls that may have been tipped with toothless beaks in some species.
Recently some of them have been recognized as also having adaptations for digging and ripping into insect nests, similar to modern anteaters, with highly specialized forelimb bones and a massively enlarged hoked claw on each hand.
And now we have another one of these digging drepanosaurs: Unguinychus onyx, whose name delightfully translates to "claw claw claw"!
Living in what is now New Mexico, USA during the late Triassic, around 215-208 million years ago, Unguinychus is only known from its enlarged hand claws but was probably similar in size to some of its close relatives, likely around 40cm long (~1'4").
Based on skin impressions from the early drepanosaur Kyrgyzsaurus it also would have been covered in small scales, possibly with a skin crest and a chameleon-like throat sac.
Drepanosaurs' evolutionary relationships are rather unclear, with various studies classifying them as an early branch of diapsid reptiles, as close relatives of the gliding kuehneosaurids, or as protorosaurian archosauromorphs. But recently another idea has been proposed, instead placing them slightly further up the archosauromorph evolutionary tree in the allokotosaur lineage close to trilophosaurids – and notably making them very closely related to fellow Triassic bird-headed weirdo Teraterpeton.
———
NixIllustration.com | Tumblr | Patreon
References:
Alifanov, V. R., and E. N. Kurochkin. "Kyrgyzsaurus bukhanchenkoi gen. et sp. nov., a new reptile from the Triassic of southwestern Kyrgyzstan." Paleontological Journal 45 (2011): 639-647. https://www.researchgate.net/publication/257843064_Kyrgyzsaurus_bukhanchenkoi_gen_et_sp_nov_a_New_Reptile_from_the_Triassic_of_Southwestern_Kyrgyzstan
Buffa, Valentin, et al. "‘Birds’ of two feathers: Avicranium renestoi and the paraphyly of bird-headed reptiles (Diapsida:‘Avicephala’)." Zoological Journal of the Linnean Society (2024): zlae050. https://doi.org/10.1093/zoolinnean/zlae050
Jenkins, Xavier A., et al. "Using manual ungual morphology to predict substrate use in the Drepanosauromorpha and the description of a new species." Journal of Vertebrate Paleontology 40.5 (2020): e1810058. https://www.researchgate.net/publication/344545876_Using_Manual_Ungual_Morphology_to_Predict_Substrate_Use_in_the_Drepanosauromorpha_and_the_Description_of_a_New_Species
Pugh, Isaac, et al. "A new drepanosauromorph (Diapsida) from East–Central New Mexico and diversity of drepanosaur morphology and ecology at the Upper Triassic Homestead Site at Garita Creek (Triassic: mid-Norian)." Journal of Vertebrate Paleontology (2024): e2363202. https://doi.org/10.1080/02724634.2024.2363202
#science illustration#paleontology#paleoart#palaeoblr#unguinychus#clawclawclaw#drepanosauridae#drepanosauromorpha#drepanosaur#allokotosauria#maybe#archosauromorpha#reptile#art#triassic weirdos#THE CLAAAWWW
262 notes
·
View notes
Video
Green-and-black Fruiteater by Jay Packer Via Flickr: Reserva Ecológica Río Blanco, Colombia
#Animalia#Amniota#Aves#Cotingidae (Cotingas)#Vertebrata#Archosauromorpha#Chordata#Reptilia#Tetrapoda#Diapsida#Colombia#Green-and-black Fruiteater#Passeriformes#South America#Animals#Birds#Pipreola riefferii#flickr
7 notes
·
View notes
Text
Mod 3 General Topics
Temporal Fosse in Reptiles
The class reptilia is divided into 5 subclasses on the bases of presence or absence certain openings through the temporal region of the skull.
Subclass I: Anapsida
Primitive reptiles with a solid skull roof. No temporal openings.
Subclass II: Euryapsida (extinct)
Skull with a single dorso-lateral temporal opening on either side, bounded by postorbital and squamosal bones.
Subclass III: Parapsida (extinct)
Skull with a single dorso-lateral temporal opening on either side bounded below by the supratemporal and postfrontal bones.
Subclass IV: Synapsida (extinct)
Skull with a single lateral temporal opening on either side bounded above by the postorbital and squamosal bones.
Subclass V: Diapsida
Skull with two temporal openings on either side separated by the bar of post orbital and squamosal bones.

Order Rhynchocephalia
Body small, elongated, and lizard like
Limbs pentadactyl, clawed, and burrowing.
Skin covered by granular scales and a mid-dorsal row of spines.
Skull diapsid. Nasal openings separate. Parietal foramen with vestigeal pineal eye present. Quadrate is fixed.
Vertebrae amphicoelous or biconcave. Numerous abdominal ribs present.
Teeth acrodont. Cloacal aperture transverse.
Heart incompletely 4-chambered
No copulatory organ in male.
Crop Milk
At the base of the neck and just in front of the sternum, the middle of the esophagus expands into a thin-walled, bilobed, elastic sac called the crop.
The crop serves as a food reservoir into which the hurriedly swallowed, dry and hard food grains are stored, moistened, and softened.
Pigeons have a unique ability to produce 'crop milk', a soft cheesy and nourishing secretion. Both sexes can produce it, especially during breeding season.
It is formed by the degeneration of the epithelial cells lining the crop.
It is regurgitated into the mouth of the young bird till they are old enough to manage a grain-diet like their parents.
Prolactin hormone, secreted by the anterior lobe of the pituitary gland, stimulates and controls the formation of crop milk.
Crop milk contains water, protein, fat, lactose, and ash.
Some parrots, flamingos, and penguins also produce crop milk
Flight Adaptations in Birds:
1. Shape: The perfectly streamline spindle-shaped body is designed to offer minimum resistance from wind, allowing it to easily through air.
2. Compact body: The light but strong dorsally and heavier ventrally body helps in maintaining balance in air. The attachment of wings high up, the high positioning of lighter organs, and the lower positioning of heavy organs and muscles all factor to give the body a low center of mass.
3. Body-covering of feathers: The smooth, closely fitting backwardly directed feathers make the body more streamlined and reduce air friction to a minimum. They also act as a blanked that envelopes air around the bird which adds to they buoyancy. The nonconducting covering of feathers insulates the body and prevents loss of heat.
4. Forelimbs modified into wings: Wings are the special flight muscles which act as instruments of propulsion through air. The particular shape of the wing, with a thick strong leading edge, convex upper surface, and concave lower surface, causes reduction in air pressure above and increase below, with minimum turbulence behind.
5. Short tail: A short muscular tail with a series of long caudal feathers arranged in a fan like manor. It serves as a rudder for steering during flight and as a counterbalance in perching.
6. Beak: The mouth is drawn out into a horny beak, it is used as a forcep. It is used for feeding, preening, nest building, offence, and defense.
7. Mobile neck and head: The neck of birds is very long and flexible. Since the beak is used for so much, mobility of the head is important.
8. Bipedal locomotion: The hindlimbs spring somewhat anteriorly from the trunk to balance and support the entire weight of the body. They are used for locomotion on ground or in water. Bipedality is characteristic of birds, their legs are relatively strong.
9. Integument: The loose skin is responsible for extensive movement of the skeletal musculature.
10. Large flight muscles: Muscles on the back are greatly reduced while the flight muscles on the breast are greatly developed, weighing nearly 1/6th of the entire bird.
automatically clamped to its perch.
11. Perching: The hind limbs of a bird are good for an aboreal life. Their leg muscles are well developed and help in perching. As the bird settles on a tree, the bending of legs exerts a pull on the flexor tendons which makes the toes automatically flex and grip the branch.
12. Endoskeleton: The fusion of the bones built with the smallest amount of material follows the Hollow-Girder principle. It combines strength with lightness, one of the first essentials in successful flight. Most of the bones are pneumatic and filled with air sacs instead of marrow. Skull bones are light and most of them are firmly fused together. Thoracic ribs are compact, necessary for flight, by concentrating the mass.
13. Digestive system: The rate of metabolism of birds is very high. The digestive system is compact but effective. The rectum is short because the faecal matter is relatively small. As they cannot afford to be weighed down with excess faecal weight, it is excreted immediately.
14. Air sacs and Respiration: The lungs have a system of air-sacs attached to them, which occupy all the available space between internal organs, even extending to the cavities of hollow bones. The air-sacs secure more perfect aeration of lungs and help in the regulation of the body temperature.
15. Warm blooded: Birds are warm-blooded. The perfect aeration of blood is responsible for the high temperature of body (40-46°C), which is necessity for flight requiring a great output of energy over a long period.
16. Circulatory system: Rapid metabolism and warm-bloodedness require a large oxygen supply and an efficient circulatory system.
17. Single ovary: Presence of a single functional ovary on the left side in the female bird leads to reduction of weight which is essential for flight.
Migration in Birds
Bird migration is a two-way journey. It is a regular, periodic, to-and-fro movement of a population of some birds between their summer and winter homes, or from a breeding and nesting place to a feeding and resting place.
Not all birds migrate, the ones that stay in one place all year round are called residential birds, ex: bobwhites
There are various types of migration, for example:
1. Latitudinal migration: Migration along the latitudinal lines of the earth, like north to south and vice versa. Ex: heading up north to avoid summer heat and moving back south to avoid cold winters.
2. Longitudinal migration: Migration along the longitudinal lines of the earth, like east to west and vice versa. Ex: moving from a breeding area in Asia to the Atlantic coast to avoid continental winter.
3. Altitudinal migration: Migration up and down the slopes of mountains to different altitudes. Ex: moving up to the top during summers and back down to the bottom during winters. Aka vertical migration.
4. Partial migration: In addition to residential birds, an influx of new birds of the same species for a short period of time. Ex: blue jays in North America traveling south to join the sedentary populations in the southern states.
5. Irregular migration: In some birds after breeding, the adults and young may stray from their home to disperse in all directions over many or a few 100 miles in search of food and safety from enemies. (??What) Ex: Herons.
6. Seasonal migration: In Britain, nightingales are considered to be summer visitors because the arrive in spring from the south, remain to breed, then leave back south in the autumn. Meanwhile redwings are considered winter visitors because they arrive in the autumn from the north, stay for the winter, and fly back north in the spring.
Modes of Flight in Migration:
1. Nocturnal and Diurnal:
Birds which migrate during the day time are called diurnal migrants. They have a greater tendency to travel in flocks, be it well organized flocks like ducks or loose flocks like swallows.
Birds which migrate during the night time are called nocturnal migrants. They fly late to avoid predators in the dark. Ex: small birds like sparrows.
2. Segregation during migration:
Some birds travel in separate companies such as kingfishers
Others travel in mixed companies of several species due to similar size or common method of feeding. Ex: swallows and blue birds.
In some species, male and female birds travel separately, males arrive first to build nests while the young usually travel with the females.
3. Range of migration:
The distance traveled by migratory birds depends upon local conditions and the species concerned.
The Himalayan snow partridge only descends a few 100 feet, covering hardly a mile or two while the chicades travel down nearly 2500 meters.
The longest traveling birds is the artic turn. It breeds in the summer far in the Artic circle and migrates a distance of 17,500km to reach the edges of Antarctica by winter, then returns the same way by summer.
4. Altitude of flight:
As in how close to the earth birds are flying at. Most migration takes place within 1000 meters of the earth. Some small nocturnal migrant birds fly at 1500 to 4000 meters.
Some species cross the Andes and the Himalayas at altitudes of 6000 meters or more.
5. Speed and duration of flight:
The average flight velocity of small birds exceeds 50kph, the greatest speed recorded in India is of two species of swifts, 250-325kph.
Several hundreds of kilometers may be covered nonstop, with an average of 800km. The golden plover holds the world record for the longest nonstop bird flight, from Hudson bay in Alaska to South America, 3850km.
6. Regularity of Migration:
Several species show an incredible regularity when it comes to their arrival and departure. Despite long distances and harsh weather, they are often very punctual when it comes to their time of arrival. Sometimes they even come back to the exact same breeding place every year.
7. Routes of Migration:
Birds usually follow definite lines of flight.
Nocturnal migration of small land birds proceeds with the general airflow on a broad front. In the spring, it occurs northwards along warm air currents from the south. In the autumn, it occurs southwards on the cool winds of the north.
Deviation in path can occurs due to configuration of land, coastline, courses of great rivers or intervening mountain chains, etc.
1 note
·
View note
Text
yes, I know wikipedia says "reptilia" is a subset of sauropsida, but that is hardly a consensus/universal agreement. Plus, this is a silly tumblr poll, and I enjoy silliness. (Including an option does not mean I endorse it). Some pros and cons are below. don't worry, I plan on doing this for other holdover linnean names folks can't let go of - fish, amphibian, bird, mammal, bug...
Pro to Reptiliomorpha: no way you're leaving anything out Con to Reptiliomorpha:
Pro to Amniota: "mammal-like reptile" stays Con to Amniota: it is absolute madness to consider mammals reptiles, that is such a deep divergence
Pro to Sauropsida: mammals are left out, as are all synapsids, but most things called reptiles stay in (as do birds) Con to Sauropsida: people will still call Dimetrodon a reptile and we will never know peace
Pro to Eureptilia: excluding parareptiles which are just weird enough that you could do it Con to Eureptilia: it's somewhat arbitrary and excludes a bunch of things we'd call reptiles today
Pro to Diapsida: being a diapsid is usually considered a major feature of reptiles, esp now we know turtles evolved from diapsids Con to Diapsida: it's somewhat arbitrary and excludes a bunch of things we'd call reptiles today
Pro to Sauria: crown group of living reptiles Con to Sauria: ichthyosaurs (maybe) & a bunch of other reptile like things are excluded
Pro to Lepidosauromorpha: birds aren't called reptiles Con to Lepidosauromorpha: crocodiles, turtles, pterosaurs, nonavian dinosaurs, and many more aren't reptiles
#reptiles#palaeoblr#taxonomy#animals#herpetofauna#I do this to sow chaos#and wait for my beta readers to get back on chapter one#and avoid doing my research#don't tell my advisor
133 notes
·
View notes