#debivort
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
Tumblr was named as a finalist in Lead411’s New York City Hot 125 in Aug 2010.
Photo

Various coquina shells. Individuals in the mollusk species Donax variabilis show diverse coloration and patterning in their phenotypes. by Debivort
7K notes
·
View notes
Text


Green tiger barb (Puntigrus tetrazona)
Source: Debivort at Wikimedia Commons
48 notes
·
View notes
Text

Various coquina shells. Individuals in the mollusk species Donax variabilis show diverse coloration and patterning in their phenotypes.
by Debivort
3 notes
·
View notes
Text

Dwarf gourami (upload by Debivort)
#sp: trichogaster lalius#ge: trichogaster#fa: osphronemidae#or: anabantiformes#cl: actinopterygii#ph: chordata#ki: animalia
1 note
·
View note
Text
Maiasaura peeblesorum
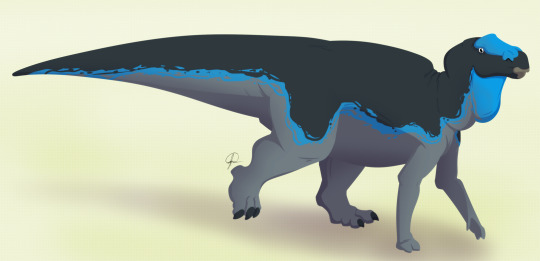
By José Carlos Cortés
Etymology: Good Mother Reptile
First Described By: Horner & Makela, 1979
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Ornithischia, Genasauria, Neornithischia, Cerapoda, Ornithopoda, Iguanodontia, Dryomorpha, Ankylopollexia, Styracosterna, Hadrosauriformes, Hadrosauroidea, Hadrosauromorpha, Hadrosauridae, Euhadrosauria, Saurolophinae, Brachylophosaurini
Status: Extinct
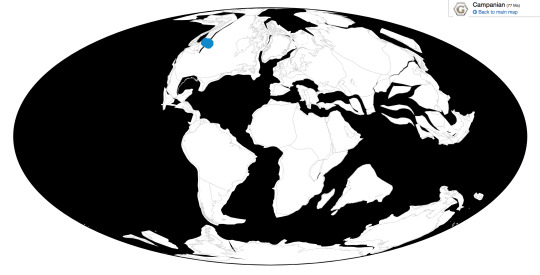
Time and Place: Between 77.2 and 76.3 million years ago, in the Campanian of the Late Cretaceous
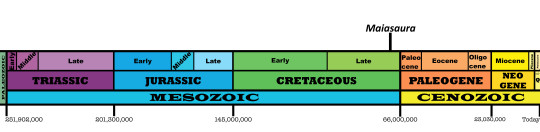
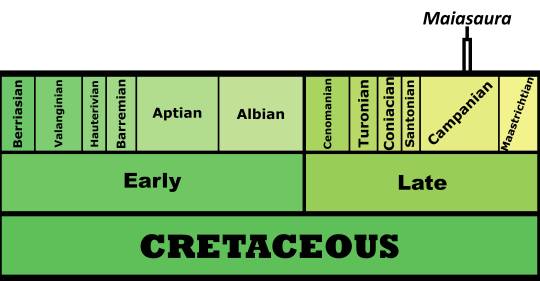
Maiasaura is known from the Upper member of the Two Medicine Formation in Montana
Physical Description: Maiasaura was a medium-sized hadrosaur - aka, a “duck-billed” dinosaur. Famed for being known from hundreds of individual skeletons, we have a general idea of the appearance of this dinosaur at every stage of its life cycle. Baby Maiasaura were around 0.4 meters in length and were positively tiny in weight, weighing less than 250 kilograms. These babies were adorable in appearance: with large eyes, small heads, and small limbs. The limbs were very weak and skinny at this point in life. Despite this extremely small size, Maiasaura young grew quickly - growing to 1.5 meters in length by the first year, and reaching sexual maturity at about the age of three or so, when they weighed around 1250 kilograms. Full skeletal maturity then came at about five years of age. At this point, Maiasaura were as much as 3000 kilograms in weight, and reaching 9 meters in length. Maiasaura adults were much beefier than the young - with thick, strong hind legs and somewhat more gracile front legs, it was almost as if they had deer front legs and elephant hind legs. The front feet formed hoof-like structures - with the pinky and thumb both sticking out, the middle three fingers were fused together. The hind limbs were typical ornithopod feet, with three toes splayed out like that of a very thick bird. Their tails were thick and muscular, and their torsos also very beefy. They had very thick, muscular necks as well.

By Ripley Cook
The heads of Maiasaura were rectangular and long, with flattened duck bills in the front. In the jaws, there were rows upon rows of densely packed teeth, forming a single surface. This surface was essentially serrated with the number of teeth packed in there. The upper jaws could then expand, allowing the lower jaws to slide upwards into them, creating a chewing motion. The more duck-like front bill was used to snip off plants and bring them into the jaw. Maiasaura also had a very large nose, forming a sort of lump in the front of the snout - this would have helped keep the head cool, and also allowed Maiasaura to make a variety of calls without a hollow crest attached. Above the eyes the skull of Maiasaura was domed with the brain area. In front of the eyes, on the top of the skull, there was a little ridged crest for display. It is logical to suppose that said crest would be somewhat patterned or even colorful, for display.
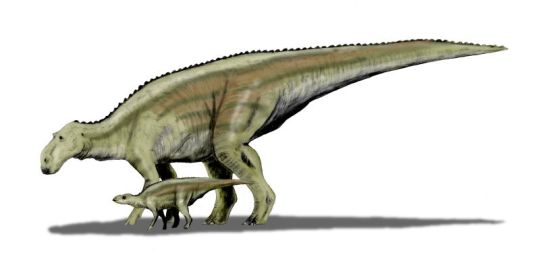
By Nobu Tamura, CC BY 3.0
Scale impressions from Maiasaura are known. Adults of this species were entirely scaley, with almost a pebble-like texture of scales covering the entire body. These small round patches didn’t seem to overlap much, but were densely packed and not leaving much in the way of bare skin showing. Very small ones no longer than 2 millimeters were interspersed with bigger, more hexagonal ones at five to ten millimeters long. It is possible that fuzz would extend between the scales, but they would have looked rather like plants growing between sidewalk scales, and fairly impossible to see ultimately. The back was bumpy from the spine, and rather high over the animal - making Maiasaura itself quite tall. The scales were even bigger on this portion of the animal. Though skin impressions are known from Maiasaura adults and close relatives, baby Maiasaura do not have preserved skin impressions. What this means is that, while it seems very logical they’d also be scaly, there is a possibility they were fluffy to stay warm, given their smaller size. We present one hypothetical reconstruction of such for you all below, with the caveat that it is purely speculation at this time.

By Diane Ramic
Diet: Maiasaura, like other hadrosaurs, fed mainly on soft, wet vegetation at low and middle levels of browsing (rather more tough, hard, dry vegetation like scrub plants and desert brush). So, it would favor leaves, berries, and more tender shoots, as well as plants in sources of water.

By Madchester, in the Public Domain
Behavior: Maiasaura was a highly social, active animal - warm blooded in energy levels, these dinosaurs would have spent most of their time, each and every day, wandering around looking for food and socializing with other members of the herd. They spent a good portion of their time taking care of their young, of course, but that was only during the breeding season. Nests were made in large breeding colonies, not unlike their modern bird relatives such as seabirds, with gaps between nests only 7 meters long - less than the length of the adults that had to move between them! Between thirty and forty eggs were laid in a dense spiral pattern, and these eggs were the size of an ostrich’s today. Rotting vegetation was placed upon the nest to keep it warm. The babies, not able to take care of themselves upon hatching, entirely relied on their parents to bring them back chewed up food and look out for their safety. Sadly, most of the young would still die in the first year of life - mainly due to disease and predation, up to 90% of the young would die in the first year of life. Still, the parents did their best - with the young having features associated with cuteness, indicating dependence on the parents for survival until they reached larger sizes.

By Debivort, CC BY-SA 3.0
Past that point, however, as the young grew faster, they fared better in terms of mortality - dropping down to 12% mortality until reaching old age again. They began to move on their own and keep up with the herd as it moved about. Young Maiasaura would walk on two legs, and as they got heavier they would switch to four, still sometimes only using the hind limbs when needed. Upon reaching sexual maturity at around three years of age, they began to get even bulkier. The Maiasaura would live in herds hundreds of individuals large, which would have been very noisy - using those bulky nostrils to make very loud, differing calls. Come mating time, they would display to each other with the ridges on their heads and other patterns. It is uncertain who was in charge of caring for the young, as sexual dimorphism isn’t seen in the skeletons of Maiasaura - if it was just the mother, both parents, just the father, or even the parents and previous children, we do not know at this time. The herd structure would protect the young, the sick, and the old from predators, and they would probably call to each other to ensure that they stayed safe in the face of predation. That being said, most of the rest of Maiasaura would then die in old age, with the death rate jumping up to 44% at the oldest ages of 12 to 14, when their own weight, slowness, and illness would leave them more vulnerable to predators.

By Fabrizio De Rossi, retrieved from Earth Archives
Ecosystem: The Two Medicine Formation was one of the most iconic dinosaur ecosystems of all time, sort of a precursor in many ways to the more famous Hell Creek, but with more variety and dinosaur diversity! Here was a very large floodplain, filled with rivers and deltas and associated plantlife on sandy riverbanks. This environment was highly associated with the ever-present Western Interior Seaway, much like the later Hell Creek. It was seasonally arid, with rainshadows from the nearby Cordilleran Highlands, which may have been at least somewhat volcanic. This made the Two Medicine Environment positively volatile - with flash flooding, droughts, dehydration, and volcanic activity all allowing for the animals in this region to be wonderfully preserved (allowing us to know so much about Maiasaura)! Plants would grow very rapidly each wet season, making the area a very lush habitat for about half the year, allowing for all these dinosaurs to congregate here. This environment was filled with conifers and pine trees primarily, though there were also other types of plants as well. There were non-dinosaurs here as well - the pterosaurs Montanazhdarcho and Piksi, the Choristodere Champsosaurus, unnamed crocodylians, lizards like Magnuviator, mammals such as Cimexomys, Paracimexomys and Alphadon, and a wide variety of turtles like Basilemys.

By Sam Stanton
Still, dinosaurs were the primary feature of the later (Upper) Two Medicine environment where Maiasaura frequented. There were four different types of Ceratopsians: the flat-nosed Achelosaurus, the curved-horned Einiosaurus, the giant-horned Rubeosaurus/Styracosaurus (depending on who you talk to, lumping-wise), and the small herbivore Prenoceratops. Ankylosaurs came in three different varieties - the large-spined but wiggle-taled Edmontonia, and the wide tail-clubbed Dyoplosaurus and Scolosaurus. Other hadrosaurs shared this environment with Maiasaura, like the large-nosed Gryposaurus, the round-crested Hypacrosaurus, and the small pointed crest having Prosaurolophus. There was also the small, active burrower Orodromeus. As for theropods, there were two different raptors - Bambiraptor and Richardoestesia - which would have been major problems for younger Maiasaura and the babies and eggs. The predatory opposite-bird, Gettyia, would have also been a predator of these smaller individauls. The troodontid Saurornitholestes would have been a major danger to these young Maiasaura along with its close cousins. The adults, on the other hand, had not one but two different species of tyrannosaur to contend with: the bulky and rarer Daspletosaurus, and the more slender Gorgosaurus that has been hypothesized to feed more on hadrosaurs than its cousin (though this is under hot debate). In short, this was the place to be to see just how diverse and fascinating non-avian dinosaurs were right at the end of their tenure, and Maiasaura was a major part (if not the most common part) of that ecosystem.

By Scott Reid
Other: Maiasaura is the closest thing non-avian dinosaurs get to a “model organism” - a creature with enough specimens, research, and data about it to use it as an example for other animals which we know less about. With hundreds of specimens found and counting, we have recorded a complete growth sequence of this dinosaur, knowing what the trajectory of a typical Maiasaura life was like. This is of vital importance, as hadrosaurs were some of the most diverse dinosaurs at the end of the Cretaceous - the end of the time of non-avian forms. It is also fascinating for how much the life history of Maiasaura - a dinosaur not close to being a bird by any stretch of the imagination - is so similar to birds. With similar rapid growth rates as their warm-blooded cousins, and similar nesting and group living strategies, Maiasaura showcases how complex behavior and lifestyles were common over the entirety of the dinosaur group. Maiasaura is also of fundamental importance because, with its discovery and descriptions in the late 1970s, it served with Deinonychus to show how dinosaurs weren’t slow, sluggish, giant lizards - but active, warm-blooded avian precursors. Dinosaurs were active, behaviorally complex, and took care of their young - something that was a truly revolutionary statement before these dinosaurs were named! So, despite not really looking like much, Maiasaura is probably one of the most important dinosaur discoveries ever found. Maiasaura itself is closely related to dinosaurs such as Brachylophosaurus, and is in general part of the “crestless” hadrosaur group, along with the contemporary Gryposaurus and the later Edmontosaurus.
~ By Meig Dickson
Sources under the Cut
Arbour, Victoria M.; Currie, Philip J. (8 May 2013). "Euoplocephalus tutus and the Diversity of Ankylosaurid Dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA". PLOS ONE. 8 (5): e62421.
Atterholt, J.; J. Howard Hutchison; Jingmai K. O’Connor (2018). "The most complete enantiornithine from North America and a phylogenetic analysis of the Avisauridae". PeerJ. 6: e5910.
Bailleul, A. M., B. K. Hall, and J. R. Horner. 2013. Secondary cartilage revealed in a non-avian dinosaur embryo. PLoS ONE 8(2):e56937:1-5
Bell, P. R. 2014. A review of hadrosaurid skin impressions. In D. A. Eberth & D. C. Evans (ed.), Hadrosaurs 572-590
Bell, P. R., R. Sissons, M. E. Burns, F. Fanti, and P. J. Currie. 2014. New saurolophine material from the upper Campanian-lower Maastrichtian Wapiti Formation, west-central Alberta. In D. A. Eberth & D. C. Evans (ed.), Hadrosaurs 174-190
Bonaparte, J. F., Novas, F. E., & Coria, R. A. (1990). Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia. Contributions in Science. Natural History Museum of Los Angeles County, 416, 1-42.
Campione, N. E., K. S. Brink, E. A. Freedman, C. T. McGarrity, and D. C. Evans. 2013. ‘Glishades ericksoni’, an indeterminate juvenile hadrosaurid from the Two Medicine Formation of Montana: implications for hadrosauroid diversity in the latest Cretaceous (Campanian-Maastrichtian) of western North America. Palaeobiodiversity and Palaeoenvironments 93:65-75
Carr, Thomas D.; J. Varricchio, David; C. Sedlmayr, Jayc; M. Roberts, Eric; R. Moore, Jason (30 March 2017). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7: 1–11.
Carroll, R. L.. 1988. Vertebrate Paleontology and Evolution 1-698
Carroll, N. REASSIGNMENT OF MONTANAZHDARCHO MINOR AS A NON-AZHDARCHID MEMBER OF THE AZHDARCHOIDEA, SVP 2015.
Chapman, R., and M. K. Brett-Surman. 1990. Morphometric observations on hadrosaurid ornithopods. In K. Carpenter and P. J. Currie (eds.), Dinosaur Systematics: Perspectives and Approaches, Cambridge University Press, Cambridge 163-177
Chinnery, Brenda J.; Horner, John R. (2007). "A New Ceratopsian Dinosaur Linking North American and Asian Taxa". Journal of Vertebrate Paleontology. 27 (3): 625–641.
Cruzado-Caballero, P., X. Pereda-Suberbiola, and J. I. Ruiz-Omeñaca. 2010. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the latest Cretaceous of Arén (Huesca, Spain). Canadian Journal of Earth Sciences 47:1507-1517
Cruzado-Caballero, P., and J. E. Powell. 2017. Bonapartesaurus rionegrensis, a new hadrosaurine dinosaur from South America: implications for phylogenetic and biogeographic relations with North America. Journal of Vertebrate Paleontology 37(2):e1289381:1-16
Cubo, Jorge; Woodward, Holly; Wolff, Ewan; Horner, John R. (2015). "First Reported Cases of Biomechanically Adaptive Bone Modeling in Non-Avian Dinosaurs". PLoS ONE. 10 (7): e0131131.
Currie, Trexler, Koppelhus, Wicks and Murphy (2005). "An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)." P.p. 313-324 in Carpenter, K. (ed.), The Carnivorous Dinosaurs. III. Theropods as living animals.
Czerkas, S. A. (1992). Discovery of dermal spines reveals a new look for sauropod dinosaurs. Geology, 20(12), 1068-1070.
D’Emic, M. D. 2015. Comment on “Evidence for mesothermy in dinosaurs.” Science 348 (6238): 982.
Davis, M. (2012). Census of dinosaur skin reveals lithology may not be the most important factor in increased preservation of hadrosaurid skin. Acta Palaeontologica Polonica, 59(3), 601-605.
Dilkes, D. W. 1999. Appendicular myology of the hadrosaurian dinosaur Maiasaura peeblesorum from the Late Cretaceous (Campanian) of Montana. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 90 (2): 87 - 125.
Dilkes, D. W. 2001. An ontogenetic perspective on locomotion in the Late Cretaceous dinosaur Maiasaura peeblesorum (Ornithischia: Hadrosauridae). Canadian Journal of Earth Sciences, 38(8), 1205-1227.
Dodson, P. 1975. Taxonomic implications of relative growth in lambeosaurine hadrosaurs. Systematic Zoology 24:37–54.
Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 116-117.
Dodson, P., C.A. Forster, and S.D. Sampson. 2004. Ceratopsidae in Weishampel, D.B., P. Dodson, and H. Osmolska (eds.) The Dinosauria. 2nd Edition, University of California Press.
Farke, A. A., D. J. Chok, A. Herrero, B. Scolieri, and S. Werning. 2013. Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids. PeerJ 1:e182.
Farlow, James O.; Pianka, Eric R. (2002). "Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs". Historical Biology. 16 (1): 21–40.
Federico L. Agnolin & David Varricchio (2012). "Systematic reinterpretation of Piksi barbarulna Varricchio, 2002 from the Two Medicine Formation (Upper Cretaceous) of Western USA (Montana) as a pterosaur rather than a bird" (PDF). Geodiversitas. 34 (4): 883–894.
Forster, C. A., and P. C. Sereno. 1994. Phylogenetic analysis of hadrosaurid dinosaurs. Journal of Vertebrate Paleontology 14(3, suppl.):25A
Forster, C. A. 1997. Hadrosauridae. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 293-299
Fowler, D. W. 2017. Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America. PLoS ONE 12 (11): e0188426.
Gates, T. A., J. R. Horner, R. R. Hanna and C. R. Nelson. 2011. New unadorned hadrosaurine hadrosaurid (Dinosauria, Ornithopoda) from the Campanian of North America. Journal of Vertebrate Paleontology 31(4):798-811
Godefroit, P., Z.-M. Dong, P. Bultynck, H. Li, and L. Feng. 1998. New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, P. R. China). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 68(supplement):3-70
Godefroit, P., V. R. Alifanov, and Y. L. Bolotsky. 2004. A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Science de la Terre 74(supplement):139-154
Godefroit, P., S. Hai, T. Yu and P. Lauters. 2008. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China. Acta Palaeontologica Polonica 53(1):47-74
Grady, J. M., B. J. Enquist, E. Dettweiler-Robinson, N. A. Wright, F. A. Smith. 2014. Evidence for mesothermy in dinosaurs. Science 344 (6189): 1268-1272
Guenther, M. F. 2009. Influence of sequence heterochrony on hadrosaurid dinosaur postcranial development. The Anatomical Record 292:1427-1441
Horner, J. R., and R. Makela. 1979. Nest of juveniles provides evidence of family structure among dinosaurs. Nature 282:296-298
Horner, J. R. 1983. Cranial osteology and morphology of the type specimen of Maiasaura peeblesorum (Ornithischia: Hadrosauridae), with discussion of its phylogenetic position. Journal of Vertebrate Paleontology 3(1):29-38
Horner, Jack and Gorman, James. (1988). Digging Dinosaurs: The Search that Unraveled the Mystery of Baby Dinosaurs, Workman Publishing Co.
Horner, J. R. 1992. Cranial morphology of Prosaurolophus (Ornithischia: Hadrosauridae) with descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships. Museum of the Rockies Occasional Paper 2:1-119
Horner, J. R., A. De Ricqles, K. Padian. 2000. Long bone histology of the hadrosaurid dinosaur Maiasaura peeblesorum: growth dynamics and physiology based on an ontogenetic series of skeletal elements. Journal of Vertebrate Paleontology 20 (1): 115 - 129.
Horner, J. R., Schmitt, J. G., Jackson, F., & Hanna, R. (2001). Bones and rocks of the Upper Cretaceous Two Medicine-Judith River clastic wedge complex, Montana. In Field trip guidebook, Society of Vertebrate Paleontology 61st Annual Meeting: Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains. Museum of the Rockies Occasional Paper (Vol. 3, pp. 3-14).
Horner, J. R., D. B. Weishampel, and C. A. Forster. 2004. Hadrosauridae. In D. B. Weishampel, H. Osmolska, and P. Dodson (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 438-463
Hunt, A. P., and S. G. Lucas. 1992. Stratigraphy, paleontology and age of the Fruitland and Kirtland formations (Upper Cretaceous), San Juan Basin, New Mexico. In S. G. Lucas, B. S. Kues, T. E. Williamson & A. P. Hunt (eds.), New Mexico Geological Society, 43rd Annual Fall Field Conference, San Juan Basin IV, Guidebook 43:217-239
Kirkland, J. I., R. Hernández-Rivera, T. A. Gates, G. S. Paul, S. J. Nesbitt, C. I. Serrano-Brañas, and J. P. Garcia-de Garza. 2006. Large hadrosaurine dinosaurs from the latest Campanian of Coahuila, Mexico. In S. G. Lucas and R. M. Sullivan (eds.), Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35:299-315
Lehman, T. M., 2001, Late Cretaceous dinosaur provinciality: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 310–328.
Lehman,T. M., S. L. Wick, and J. R. Wagner. 2016. Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas. Journal of Paleontology 90(2):333-356
McFeeters, B., Evans, D., & Maddin, H. (2018, May). Variation in the braincase and cranial ornamentation of Maiasaura peeblesorum (Ornithischia, Hadrosauridae) from the Campanian Two Medicine Formation of Montana: implications for brachylophosaurin ontogeny and evolution. In 6th Annual Meeting Canadian Society of Vertebrate Palaeontology May 14-16, 2018 Ottawa, Ontario (p. 37).
McGarrity, C. T.; Campione, N. E.; Evans, D. C. (2013). "Cranial anatomy and variation in Prosaurolophus maximus (Dinosauria: Hadrosauridae)". Zoological Journal of the Linnean Society. 167 (4): 531–568.
Morris, W. J. 1970. Hadrosaurian dinosaurs bills–morphology and function. Los Angeles County Museum Contributions in Science 193:1–14.
Osborn, H. F. (1912). Integument of the iguanodont dinosaur Trachodon. Memoirs of the American Museum of Natural History v. 1
Palmer, D., ed. (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. p. 148.
Penkalski, P. (2013). "A new ankylosaurid from the late Cretaceous Two Medicine Formation of Montana, USA". Acta Palaeontologica Polonica.
Pereda-Suberbiola, X., J. I. Canudo, P. Cruzado-Caballero, J. L. Barco, N. López-Martínez, O. Oms, and J. I. Ruiz-Omeñaca. 2009. The last hadrosaurid dinosaurs of Europe: a new lambeosaurine from the uppermost Cretaceous of Aren (Huesca, Spain). Comptes Rendus Palevol 8:559-572
Prieto-Márques, A., R. Gaete, G. Rivas Galobart and M. Boada. 2006. Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et. sp. nov. Journal of Vertebrate Paleontology 26(4):929-943
Prieto-Márquez, A. 2010. Glishades ericksoni, a new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous of North America. Zootaxa 2452:1-17
Prieto-Marquez, A., and G. C. Salinas. 2010. A re-evaluation of Secernosaurus koerneri and Kritosaurus asutralis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. Journal of Vertebrate Paleontology 30(3):813-837
Prieto-Márquez, A., and J. R. Wagner. 2010. Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia. Cretaceous Research 30:1238-1246
Prieto-Marquez, A. 2013. Skeletal morphology of Kritosaurus navajovius(Dinosauria: Hadrosauridae) from the Late Cretaceous of the North American south-west, with an evaluation of the phylogenetic systematics and biogeography of Kritosaurini. Journal of Systematic Palaeontology
Prieto-Márquez, A.; Wagner, J.R. (2013). "A new species of saurolophine hadrosaurid dinosaur from the Late Cretaceous of the Pacific coast of North America". Acta Palaeontologica Polonica. 58 (2): 255 - 268.
Prieto-Marquez, A., & Guenther, M. F. (2018). Perinatal specimens of Maiasaura from the Upper Cretaceous of Montana (USA): insights into the early ontogeny of saurolophine hadrosaurid dinosaurs. PeerJ, 6, e4734.
Ramírez-Velasco, A. A., M. Benammi, A. Prieto-Marquez, J. Alvarado Ortega, and R. Hernández-Rivera. 2012. Huehuecanauhtlus tiquichensis, a new hadrosauroid dinosaur (Ornithischia: Ornithopoda) from the Santonian (Late Cretaceous) of Michoacán, Mexico. Canadian Journal of Earth Sciences 49:379-395
Rogers, R.R. (1990). "Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-related mortality". PALAIOS. 5 (5): 394–413.
Russell, D. A. (1970). "Tyrannosaurs from the Late Cretaceous of western Canada". National Museum of Natural Sciences Publications in Paleontology. 1: 1–34.
Russell, D. A. 1984. A check list of the families and genera of North American dinosaurs. Syllogeus 53:1-35
Sullivan, R. M.; Lucas, S. G. (2006). "The Kirtlandian land-vertebrate "age"–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America". New Mexico Museum of Natural History and Science Bulletin. 35: 7–29.
Tan, Q. W., H. Xing, Y.-G. Hu, L. Tan, and X. Xing. 2015. New hadrosauroid material from the Upper Cretaceous Majiacun Formation of Hubei Province, central China. Vertebrata PalAsiatica 53(3):245-264
Trexler, D., 2001, Two Medicine Formation, Montana: geology and fauna: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 298–309.
Varricchio, D.J. (1995). "Taphonomy of Jack's Birthday Site, a diverse dinosaur bonebed from the Upper Cretaceous Two Medicine Formation of Montana". Palaeogeography, Palaeoclimatology, Palaeoecology. 114 (2–4): 297–323.
Varricchio, D. J. 2001. Late Cretaceous oviraptorosaur (Theropoda) dinosaurs from Montana. pp. 42–57 in D. H. Tanke and K. Carpenter (eds.), Mesozoic Vertebrate Life. Indiana University Press, Indianapolis, Indiana.
Versluys, J. 1923. Der Schädel des Skelettes von Trachodon annectens im Senckenberg-Museum. Abhandlungen Der Senckenbergischen Naturforschenden Gesellschaft 38:1–19.
Weishampel, D. B., and J. B. Weishampel. 1983. Annotated localities of ornithopod dinosaurs: implications to Mesozoic paleobiogeography. The Mosasaur 1:43-87
Weishampel, D. B., and J. R. Horner. 1990. Hadrosauridae. In D. B. Weishampel, H. Osmolska, and P. Dodson (eds.), The Dinosauria. University of California Press, Berkeley 534-561
Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. 861 pp.
Woodward, H. N., E. A. Freedman Fowler, J. O. Farlow, J. R. Horner. 2015. Maiasaura, a model organism for extinct vertebrate population biology: a large sample statistical assessment of growth dynamics and survivorship. Paleobiology 41 (4): 503-527.
Woodward, H. N. (2019). Maiasaura (Dinosauria: Hadrosauridae) Tibia Osteohistology Reveals Non-Annual Cortical Vascular Rings in Young of the Year. Frontiers in Earth Science, 7, 50.
#Maiasaura peeblesorum#Maiasaura#Dinosaur#Hadrosaur#Prehistoric Life#Paleontology#Prehistory#factfile#Saurolophine#Cretaceous#Mesozoic Monday#Palaeoblr#Dinosaurs#North America#Herbivore#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
360 notes
·
View notes
Text
12 Amazing Facts About The Last Judgement

It's one of his most famous works of art, located in one of the most magical places in the world. In this post you'll discover the ultimate list of facts about The Last Judgement, one of the ultimate masterpieces by Michelangelo, one of the most famous artists of the Renaissance. Interesting facts about The Last Judgement by Michelangelo 1. The Last Judgement is a huge fresco The Last Judgement is located in the Sistine Chapel in Vatican City, a chapel in the Apostolic Palace, the official residence of the Pope. It's also the final attraction in the Vatican Museum, one of the largest public art museums in the world. And what an attraction it is! The Last Judgement Fresco which has dimensions of 13.7 × 12 meters (539.3 × 472.4 inches) and covers the entire altar wall of the chapel! The fresco above the altar / Debivort / https://creativecommons.org/licenses/by-sa/3.0/ 2. It took Michelangelo 5 years to complete it With this knowledge, it's easy to conclude that he didn't paint this immense fresco overnight. The wall had been prepared in the year 1535 and he started painting in 1536. The original idea of the work was already conceived in 1533 though, but Michelangelo was mainly a sculptor and only reluctantly accepted the commission. Another obstacle was the existing art, because an altarpiece of the Assumption of Mary by Pietro Perugino already decorated the wall above the altar. This as eventually removed to be replaced by The Last Judgement. Read the full article
#Art#FactsabouttheLastJudgement#History#Michelangelo#Michelangelofamousworks#paintings#TheLastJudgement
0 notes
Photo



Looking for a Plan B for Planet Earth?
On October 6, 1995, astronomers Didier Queloz and Michel Mayor made an announcement that while it had been long anticipated was still stunning: they had discovered the very first exoplanet orbiting around a main sequence star, 51 Pegasi. Although three other exoplanets (one orbiting gamma Cephei, discovered in 1988 by the Canadian astronomers Bruce Campbell, G. A. H. Walker, and Stephenson Yang, confirmed in 2003; and two planets orbiting PSR 1257-12 discovered by radio astronomers Aleksander Wolszczan and Dale Frail in 1992) had been discovered prior to Mayor and Queloz’s announcement, 51 Pegasi was the first exoplanet to be discovered orbiting a main sequence star that is very much like our sun, and therefore thought capable of supporting life. The official name of this exoplanet is 51 Pegasi b, the lower case b denoting that the star has a companion that is not also a star (as in binary systems), and also referring to its unofficial name of Bellerophon. Bellerophon was the name of the hero from Ancient Greek mythology who tamed the Pegasus. It has since been noted that this planet orbits its star very close and probably has surface temperatures in excess of 1200 degrees, making it unfit for most of the life as we know it on earth.
The word exoplanet is a combination of the Ancient Greek root word prefix exo- meaning outside and the word planet. The word planet entered English as a scientific term from the Old French word planète, which came in turn from Late Latin planeta, which was itself a borrowing from the Ancient Greek (asteres) planetai meaning wandering (stars), from the verb planasthai meaning to wander. It took its modern meaning around 1630.
Constellation map courtesy Torsten Bronger. Artist’s concept of 51 Pegasi b courtesy Debivort. Image of 51 Pegasi courtesy NASA, all used with permission under a Creative Commons 3.0 license.
43 notes
·
View notes
Text
Tweeted
Delighted to share a new paper from our group: @Matt_AYSmith and Kyle Honegger, along with @MAChurgin and Glenn Turner, characterized individuality in fly odor preference, circuits regulating it, and showed that odors are represented idiosyncratically.https://t.co/RK15dbFrd7
— Benjamin de Bivort (@debivort) August 29, 2019
0 notes
Text
Truth Can Work Your Way
Truth Can Work Your Way
(click on the title to enable comment space and
“Music” is a Link to Christian Music – images from pixabay.com
share buttons at the bottom)
What is truth? Canofworms1 – Attribution: Debivort at en.wikipedia
This does open a can of worms. Right?
But, once there was only The Truth GOD offered to humans; that changed when Adam and Eve believed Satan’s lie in the Garden of Eden. It has been a…
View On WordPress
0 notes
Text
Tweeted
Isogenic lab animals behave differently. Maybe this variability arises in microenvironmental differences (rather than, e.g., molecular stochasticity). @jamilla_az, Sandra Ho, and @chelseajoleary used enrichment to test this idea in our new preprint.https://t.co/nCOayrgJRq pic.twitter.com/yxIQfscbil
— Benjamin de Bivort (@debivort) February 22, 2019
0 notes