#tetraloop
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
Tumblr has 16.74 million mobile monthly users in the US.
Text

In the raiA ncRNA structure that we recently determined, we identified a junction strand (B) that mimics both the GAAA-tetraloop (A) AND the D-loop/T-loop interaction motif (C) - how cool is that?
Find out more: https://doi.org/10.1016/j.jmb.2024.168833
#rna#research#science#structure#structuralbiology#cryoEM#junction#tetraloop#strand#motif#Dloop#Tloop#GAAA#GNRA#raiA
0 notes
Text
Double-stemmed and split structural variants of fluorescent #RNA Mango #aptamers [Article]
#aptamers with fluorogenic ligands are emerging as useful tools to quantify and track RNA molecules. The RNA Mango family of #aptamers have a useful combination of tight ligand binding, bright fluorescence and small size. However, the simple structure of these #aptamers, with a single base-paired stem capped by a G-quadruplex, can limit the sequence and structural modifications needed for many use-inspired designs. Here we report new structural variants of RNA Mango that have two base-paired stems attached to the quadruplex. Fluorescence saturation analysis of one of the double-stemmed constructs showed a maximum fluorescence that is ~75% brighter than the original single-stemmed Mango I. A small number of mutations to nucleotides in the tetraloop-like linker of the second stem were subsequently analyzed. The effect of these mutations on the affinity and fluorescence suggested that the nucleobases of the second linker do not directly interact with the fluorogenic ligand (TO1-biotin), but may instead induce higher fluorescence by indirectly altering the ligand properties in the bound state. The effects of the mutations in this second tetraloop-like linker indicate the potential of this second stem for rationale design and reselection experiments. Additionally, we demonstrated that a bimolecular mango designed by splitting the double-stemmed Mango can function when two RNA molecules are co-transcribed from different DNA templates in a single in vitro transcription. This bimolecular Mango has potential application in detecting RNA-RNA interactions. Together, these constructs expand the designability of the Mango #aptamers to facilitate future applications of RNA imaging. http://rnajournal.cshlp.org/cgi/content/short/rna.079651.123v1?rss=1&utm_source=dlvr.it&utm_medium=tumblr
0 notes
Text
Hiding in Plain Sight
Gaya Amarasinghe Washington University in St. Louis
On a recent visit to the laboratory where he worked as an undergraduate, Gaya Amarasinghe thought about his scientific journey from student to professor. At first glance, his research interests seem to have drastically changed. Back then, he was studying signaling in cancer biology, and how oncogenes communicate within cells. Now, his group at Washington University in St. Louis investigates how some of the world’s deadliest viruses, such as Ebola, outcompete the host and cause disease.
Yet, he decided, not much has changed after all. “In the last 20 years, I haven��t done anything differently,” joked Amarasinghe, who trained as a structural biologist and biochemist. “My group now studies proteins from viruses and bacteria, but they interact with and sometimes hijack the same immune signaling cascades I studied before. I’m asking the same questions.”
Amarasinghe’s group examines how the host immune system recognizes foreign pathogens and how normal immune signaling changes in the presence of microbes.

“We ask simple questions, and we answer them using simple experiments,” Amarasinghe says. “What are the initial components in the system, what does the structure look like, and what are the dynamics of these molecules? Then we ask what happens to those properties when they interact with a virus. What does that interaction look like? How does it move? What changes?”
It was a question that first made them curious about Ebola and related Marburg viruses: How do they evade the pathogen detection system that the immune system uses to distinguish self from non-self?
Most people know these filoviruses as causes of severe, often fatal, outbreaks of hemorrhagic illness, usually in Africa, where people first catch the contagious diseases from animals.
Few viruses are as dependent upon first interactions with the host as Ebola and its cousins. Unlike other viruses with multiple ways of manipulating an immune response during an infection, Ebola and Marburg viruses begin with a knock-out punch to the immune system defenses. It happens at the earliest stage, Amarasinghe says.
The critical moment comes when the innate immune cells that function as sentinel cells in the host, such as dendritic cells and macrophages, recognize the viruses as unwanted foreign invaders—or not, as is often the case.
A key player is a crucial viral protein called VP35. Without a working VP35, the viruses cannot defend early immune responses during infection. It also blocks many steps of the host’s innate immune response, including one that ramps up the antiviral immune fight.
The first attempts to answer the question about initial recognition of the virus were anything but simple. “The hardest part was getting the material we needed,” Amarasinghe says. It was a brute force effort to purify the protein. He estimates it took the equivalent of 10-20 people-years of work in many groups, including his own, to finally isolate well-behaved VP35 by genetic recombination techniques.
In 2009, his group published the first of a half-dozen papers, which include at least 10 different VP35 structures of what the interferon inhibitory domain (or IID) from several Ebola strains and Marburg, which have different disease courses.
“VP35 IID was nothing like anyone predicted,” Amarasinghe says. “This protein is hiding the non-self signal that would otherwise get recognized by the immune system. It’s hiding the signal in plain sight.” The protein doesn’t even need to move much. It has a fold that fits neatly over the non-self signal, an end of the detection RNA found in every cell.
The group also found a fundamental difference between Ebola and Marburg viruses in how they recognize the host RNA and hide it. “They are equally virulent and pathogenic, but there is a key difference at an early stage,” he says. “Why does that matter? What happens in early stages determines the outcome of infections.” For example, Reston Ebola virus, a close cousin of the more virulent filoviruses also uses VP35 to bind RNA, but differences in other parts of the virus replication cycle make Reston non-pathogenic in humans.
Once they knew what the protein looked like in the related hemorrhagic viruses, Amarasinghe’s team helped design mutated versions to model the disease and better define the biology. The mutated viruses were immunogenic.
“Very cool, because now we have virulent and avirulent near-isogenic viruses to study the disease in animal models” he says. This knowledge can be used to develop actual vaccine candidates with different molecular backbones or using smaller subunits for safety, he says.
Most of the lab’s work happens in test tubes with non-infectious viral proteins. “We have collaborators working in biosafety level 4 that put some of these crazy ideas to work,” he says. Amarasinghe and his collaborators are testing the biological mechanisms in animal models.
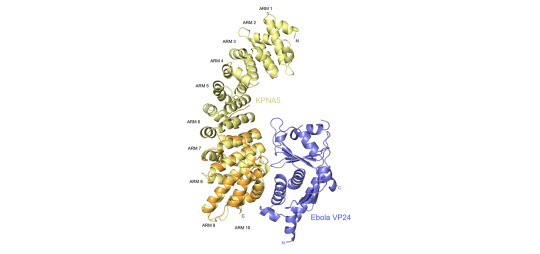
A second set of discoveries from their work helps explain why Ebola and Marburg are not responsive to interferon, a potential treatment given for many viral infections, but as it turns out it does not work well in filoviral infections. VP24, another protein encoded by the Ebola virus binds to a key cargo transporter called karyopherin, which is responsible for moving important signals, or transcription factors, into the nucleus to trigger an immune response. Viruses are known to block dendritic cells from maturing, preventing the normal interferon signaling necessary for an adaptive immune response. Amarasinghe and his colleagues have shown that VP24 also inhibits molecular signaling from therapeutically delivered interferon.
“Typically we think of a host-microbe arms race, with the host evolving away from the pathogen,” Amarasinghe says. “In this case, the place where the viral protein binds is unable to evolve away, because the host would become non responsive to its own immune signal. The virus identified an area where mutation change rate is low.”
Most of the lab projects fall into one of two categories—how host factors recognize the signature or foreign pathogens, and how host signaling changes in the presence of a microbe.
The host stress response to a virus, and not the virus itself, could be responsible for some diseases and point the way to better treatments. This could be the case with the human respiratory syncytial virus (hRSV), an infection most kids acquire by age 2. It can be fatal to infants and the elderly. It is in the same family as another emerging infectious disease people catch from animals is Nipah virus, an often fatal infection that causes encephalitis and severe respiratory distress.
Amarasinghe and his colleagues purified and solved the first crystal structure of an hRSV protein known as NS1. They reported their findings in 2017 in Nature Microbiology. They found a duplicated structural fold that suggests NS1 may have a greater role in regulating host responses than previously appreciated. “It’s not quite falling off log, but it’s close,” he says about the functional revelations.
In the future, Amarasinghe says his lab will continue to explore what drives infection in the early stages from host response point of view, aiming to understand factors that determine outcome in longer term persistent infections. The knowledge may be useful in other bacterial and viral systems, including the effect of co-infections on outcomes. “None of these exist in isolation,” he says.
Amarasinghe starts many days as early as 4:00 am and most mornings with a long run or a bike ride. It keeps him in shape for occasional long-distance mountain biking or skiing.
By his name alone, Amarasinghe may have been destined to work with proteins and RNA. Every letter in his full name represents an amino acid. He pointed out that his first name, GAYA, also forms a tetraloop (guanine-adenine-pYrimidine-adenine).
-Carol Cruzan Morton
2 notes
·
View notes
Text
Blind prediction of noncanonical RNA structure at atomic accuracy [NEW RESULTS]
Prediction of RNA structure from nucleotide sequence remains an unsolved grand challenge of biochemistry and requires distinct concepts from protein structure prediction. We report a stepwise Monte Carlo method that has enabled the first blind prediction recovering all noncanonical base pairs of a complex RNA structure, a double pseudoknot from the Zika virus genome posed as a community-wide RNA Puzzle. A benchmark of 82 diverse motif structure challenges and prospective experimental tests for three previously unsolved tetraloop/receptors support the method's general ability to recover noncanonical pairs ab initio, with remaining problems traced to limitations of current macromolecule free energy functions. — bioRxiv : Biochemistry
#bioRxiv : Biochemistry#Blind prediction of noncanonical RNA structure at atomic accuracy [NEW RESUL
0 notes
Text
PCBP2 enables the cadicivirus IRES to exploit the function of a conserved GRNA tetraloop to enhance ribosomal initiation complex formation.
Pubmed: http://dlvr.it/PKW9xf
0 notes
Text
Concentration-dependent and configuration-dependent interactions of monovalent ions with an #RNA tetraloop.
Related Articles Concentration-dependent and configuration-dependent interactions of monovalent ions with an #RNA tetraloop. J Chem Phys. 2018 Jun 14;148(22):222837 Authors: Miner JC, García AE Abstract Monovalent salt solutions have strongly coupled interactions with biopolymers, from large polyelectrolytes to small #RNA oligomers. High salt concentrations have been known to induce transitions in the structure of #RNA, producing non-canonical configurations and even driving #RNA to precipitate out of solution. Using all-atom molecular dynamics simulations, we model a monovalent salt species (KCL) at high concentrations (0.1-3m) and calculate the equilibrium distributions of water and ions around a small tetraloop-forming #RNA oligomer in a variety of structural arrangements: folded A-#RNA (canonical) and Z-#RNA (non-canonical) tetraloops and unfolded configurations. From these data, we calculate the ion preferential binding coefficients and Donnan coefficients for the #RNA oligomer as a function of concentration and structure. We find that cation accumulation is highest around non-canonical Z-#RNA configurations at concentrations below 0.5m, while unfolded configurations accumulate the most co-ions in all concentrations. By contrast, canonical A-#RNA structures consistently show the lowest accumulations for all ion species. Water distributions vary markedly with #RNA configuration but show little dependency on KCL concentration. Based on Donnan coefficient calculations, the net charge of the solution at the surface of the #RNA decreases linearly as a function of salt concentration and becomes net-neutral near 2.5-3m KCL for folded configurations, while unfolded configurations still show a positive solution charge. Our findings show that all-atom molecular dynamics can describe the equilibrium distributions of monovalent salt in the presence of small #RNA oligomers at KCL concentrations where ion correlation effects become important. Furthermore, these results provide valuable insights into the distributions of water and ions near the #RNA oligomer surface as a function of structural configuration. PMID: 29907048 [PubMed - indexed for MEDLINE] http://bit.ly/2NubauY #Pubmed
0 notes
Text
Understanding In-line Probing Experiments by Modeling Cleavage of Non-reactive #RNA Nucleotides [Article]
Ribonucleic acid (RNA) is involved in many regulatory and catalytic processes in the cell. The function of any RNA molecule is intimately related with its structure. In-line probing experiments provide valuable structural datasets for a variety of RNAs and are used to characterize conformational changes in riboswitches. However, the structural determinants that lead to differential reactivities in unpaired nucleotides have not been investigated yet. In this work we used a combination of theoretical approaches, i.e., classical molecular dynamics simulations, multiscale quantum mechanical/molecular mechanical calculations, and enhanced sampling techniques in order to compute and interpret the differential reactivity of individual residues in several RNA motifs including members of the most important GNRA and UNCG tetraloop families. Simulations on the multi ns timescale are required to converge the related free-energy landscapes. The results for uGAAAg and cUUCGg tetraloops and double helices are compared with available data from in-line probing experiments and show that the introduced technique is able to distinguish between nucleotides of the uGAAAg tetraloop based on their structural predispositions towards phosphodiester backbone cleavage. For the cUUCGg tetraloop, more advanced ab initio calculations would be required. This study is the first attempt to computationally classify chemical probing experiments and paves the way for an identification of tertiary structures based on the measured reactivity of non-reactive nucleotides. http://bit.ly/2ln6yv4 #RNAJ
0 notes
Text
Revisiting GNRA and UNCG folds: U-turns versus Z-turns in RNA hairpin loops [LETTER TO THE EDITOR]
When thinking about RNA three-dimensional structures, coming across GNRA and UNCG tetraloops is perceived as a boon since their folds have been extensively described. Nevertheless, analyzing loop conformations within RNA and RNP structures led us to uncover several instances of GNRA and UNCG loops that do not fold as expected. We noticed that when a GNRA does not assume its "natural" fold, it adopts the one we typically associate with a UNCG sequence. The same folding interconversion may occur for loops with UNCG sequences, for instance within tRNA anticodon loops. Hence, we show that some structured tetranucleotide sequences starting with G or U can adopt either of these folds. The underlying structural basis that defines these two fold types is the mutually exclusive stacking of a backbone oxygen on either the first (in GNRA) or the last nucleobase (in UNCG), generating an oxygen– contact. We thereby propose to refrain from using sequences to distinguish between loop conformations. Instead, we suggest using descriptors such as U-turn (for "GNRA-type" folds) and a newly described Z-turn (for "UNCG-type" folds). Because tetraloops adopt for the largest part only two (inter)convertible turns, we are better able to interpret from a structural perspective loop interchangeability occurring in ribosomes and viral RNA. In this respect, we propose a general view on the inclination for a given sequence to adopt (or not) a specific fold. We also suggest how long-noncoding RNAs may adopt discrete but transient structures, which are therefore hard to predict.
— RNA
#RNA#Revisiting GNRA and UNCG folds: U-turns versus Z-turns in RNA hairpin loops [LETTER TO THE EDIT
0 notes