#abelisaurs are the best theropods
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
US Tumblr user growth rate is estimated to slow down to 4.1%.
Text
Showa Godzilla Biological Analysis
(including Minilla)
This Godzilla appeared in the ocean after the first godzilla died from the oxygen destroyer presumably also awakened by the american hydrogen bomb test in South pacific in 1954.
Godzilla's physiology appears similar to theropods except more like outdated depictions of theropods standing upright with an erect back and dragging its tail on the ground!
Godzilla also has large well developped arms with opposable thumbs to manipulate his environment and also grab onto opponents.
Godzilla's tissues are theorized to be so dense that they’re stronger than the strongest materials in universe as he was durable enough to tank the space monster King Ghidorah's gravity beams which are stated to be powerful enough to destroy planets like venus, this gives him incredible resilience!
Godzilla is also shown to be quite adaptable as electricity used to harm him but now became immune to it even absorbing soo much lightning that he gained electromagnetism during his battle against Mechagodzilla as he uses it to attract the robot to him as it was attempting to retreat!
Godzilla is shown to be incredibly intelligent (like with most showa kaijus) capable of having long term memory and also showing alot of emotions as seen with Minilla showing sympathy for him and adopts him and also shows compassion for him caring and teaching him how to use an atomic breath, he does show however to be extremely aggressive towards humans in the past but it was stated by the shobijins that he hates humans because they hate them, probably due to being obviously threatened by the millitary who shot at him when he was first sighted, nowadays he seems to be more gentle towars humans even saving humanity on occasion from threats that dare to threatnened earth with the help of other earth monsters!
Another godzilla individual was seen in the form of an infant godzilla named Minilla which gives us some info on the growth stages of the species!
When hatched out of the egg from a group of large praying mantis Kamacuras, Minilla possesses a tan color on his skin has a shorter neck and thinner limbs and lacks any dorsal plates, Minilla then grew to 18 meters tall his coloration became grey and possesses a more elongated head and his dorsal plates started to grow, and developped an atomic breath although unlike the mature godzilla minilla can only breathe a smoke ring possibly due to not having a well developped organ!
Most People (atleast from what i can find-) seem to think showa goji is a theropod possibly a late ceratosaur that splits off from abelisaurs but i like to think Showa Godzilla is actually a highly derived Notosuchian related to simosuchus!
This kaiju biological analysis was inspired by @iamthekaijuking's biological analysis on showa gamera kaijus i had alot of fun reading them and i also had alot of fun doing this analysis on showa goji and this is the best i can do with my research!
17 notes
·
View notes
Text
Wednesday 13/10/21 - Dinosaurs of the World Part 4; Madagascar and India

Majungasaurus grabs the tail of a Madagascan crocodilian, Mahajungasuchus; Sergey Krasovskiy
When you think of our current geography, it makes sense to think of Madagascar as part of Africa's greater biosphere; a lot of biota found there now have relatives on the African mainland. The same for India, even more so because the Indian continental plate has made landfall on the Asian Mainland. But in the Mesozoic Period, these two landmasses were more associated with the southern continents of Australia, Antarctica, and South America. India wasn't even a part of Asia until after the dinosaurs went extinct.

Late Cretaceous Madagascar and India, Walter Myers
During the age of reptiles, India and Madagascar were best buddies, and biogeographically, palaeontologists unite these two as their own distinct region. In terms of dinosaur diversity, the two regions share a lot in common with South America. Titanosaur Sauropods and Abelisaur Theropods are king in these regions. So when I started to construct a list of dinosaur highlights, I found it difficult to make a list of dinosaurs that weren't all Sauropods and Abelisaurs. So compared to my Africa, and Asia episodes, this list will be a bit shorter, but rest assured, there is a lot to discover about dinosaurs from Madagascar and India.
Isisaurus colberti

Isisaurus, Dmitry Bogdanov
Isisaurus was a large sauropod from Late Cretaceous India. It was a member of the Titanosaur clade, the largest and longest, and heaviest of all dinosaurs. Isisaurus colberti was originally called Titanosaurus colberti, united with the Indian dinosaur that this clade is named after, but closer inspection of its fossils led palaeontologists to classify Isisaurus in its own genus. Titanosaurus itself is currently a nomen dubium: a species that may not be real because remains are too fragmentary to be sure.
The most distinguishing feature of Isisaurus is its tall neck spines, giving it a very unique appearance for a Titanosaur, but it's not really known why they had them. Isisaurus coprolites (fossil poo) have had parts of pathogenic (diseasing) fungus found in them, giving paleobotanists a unique insight into the plant life of its habitat. Isisaurus is also unique in its name. It does not in fact call reference to the Egyptian God Isis, but it is fact named for the ISI (Indian Statistical Institution), making the only dinosaur I personally know to be named after an acronym.
Barapasaurus tagorei
Barapasaurus, SpinoInWonderland (deviantart)
Barapasaurus was an Early Jurassic Sauropod from central India. It was very early for a Sauropod, evolutionarily, but quite large for its time, 5.5 m tall and 14 m long. It's skeleton reveals a lot of traits basal to the true sauropod group, but was primitive in others, particularly its feet. Barapasaurus was the first dinosaur skeleton to be mounted in an Indian Museum, in 1977. It's name is also part Indian (Bengali), Bara meaning "Large", and Pa meaning "Leg" and of course the Greek Saurus to end it out. The large leg bone in question being the first bone of the animal found.
Dravidosaurus blandfordi

Dravidosaurus, Pavel Riha
Dravidosaurus was (potentially) a late surviving Stegosaur found in Late Cretaceous India. So far in these lists, I've tried to avoid highlighting Nomen Dubium, but I wanted a bit of variety in my Indian Dinosaurs, so this is an exception. Dravidosaurus is named for the Dravidanadu region of South India where it was found. Studies of the specimen in the 1990s hypothesised it was a plesiosaur, a type of marine reptiles related to turtles. But studies in the 2010s decided that among the remains found are a Stegosaur back plate and tail spike, but there's still too little material to be sure of anything. If it was indeed a Stegosaur, it was one of the smallest, 3 metres long and maybe half a metre tall.
Rahonavis ostromi

Rahonavis, Julio Lacerda (@paleoart on tumblr)
Rahonavis was a small theropod dinosaur from Late Cretaceous Madagascar. It's exact classification has been subject to much debate since it was first described, as it lies right on the boundary between non-avian dinosaurs and the earliest true birds. Depending on who you ask, Rahonavis is either a basal Avialan like Archaeopteryx, or an extremely derived dromeosaur like Dakotaraptor or Microraptor. Given the shape of its arms, and the evidence of flight feathers, Rahonavis was likely capable of flight, but wasn't as good in the air as modern birds. It was the size of a typical modern bird too, about 70 cm long. It's name consists of the Malagasy Rahona, meaning "Cloud/Menacing" and the Latin Avis, meaning "Bird".
Majungasaurus crenatissimus

Majungasaurus, Moonmelo (deviantart)
Majungasaurus was a medium sized Abelisaur theropod from Madagascar. It was from the very end of the Cretaceous period, one of the last dinosaurs. Majungasaurus was about 3 m tall and 8 m long. It had one of the most complete skull fossils of its family, with a blunt snout, sturdy, but wrinkled face structure, and its signature cone shaped horn right between its eyes. The horn was not built for combat and would've likely been a display structure. Its short but robust jaw allowed Majungasaurus and other Abelisaurs to bite down and twist more effectively than other theropods, and may have been specialised for hunting the many Sauropod species that inhabited Madagascar in its time. Majungasaurus is named for the Mahajunga province of North Madagascar.
The unique horn shape may be the most visual diagnostic feature of Majungasaurus, but many actually know the dinosaur for being the only non-avian dinosaur to have evidence of cannibalism. Signature bite marks found in Majungasaurus prey has also been found in other adult and juvenile Majungasaurus specimens. Palaeontologists are unsure if this is evidence of Majungasaurus actively hunting other members of its species, or if they simply scavenged on the dead.
Thanks for Reading
I apologise for making this article a bit shorter than the last couple has been. I've been trying to include some variety in dinosaur clades in these regional highlights, so rather than discuss mostly Abelisaurs and Titanosaurs, I picked one of each and then a few more notable species I could find. There really are a lot of interesting species from both India and Madagascar, so I recommend looking up info on the other interesting dinosaurs from the regions.
Next blogpost I'm thinking Europe, and there's quite a few interesting species I can think of already without consulting the entire list of species. My intention is to do North America last, since most of the species people know are from there and I want my readers to savour what the rest of the world has to offer first. As usual, links to previous and future articles below, and will be edited once I have written more.
<< Part 3: Africa || Part 5: Europe >>
#blog#blogpost#palaeontology#dinosaurs#indian dinosaurs#madagascan dinosaurs#isisaurus#barapasaurus#dravidosaurus#rahonavis#majungasaurus
37 notes
·
View notes
Text
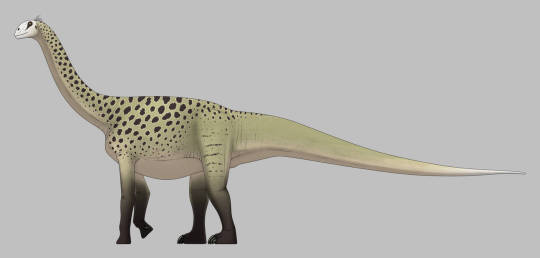
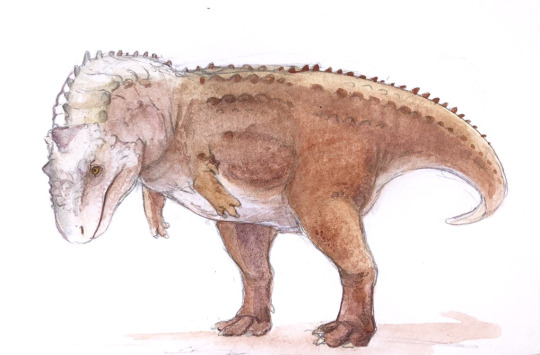
Rajasaurus narmadensis
By Ripley Cook
Etymology: King Reptile
First Described By: Wilson et al., 2003
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Theropoda, Neotheropoda, Averostra, Ceratosauria, Neoceratosauria, Abelisauroidea, Abelisauridae, Majungasaurinae
Status: Extinct
Time and Place: Between 70 and 66 million years ago, in the Maastrichtian of the Late Cretaceous
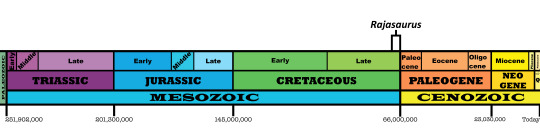

Rajasaurus is known from the Lameta Formation of Gujarat, India
Physical Description: Rajasaurus was an Abelisaur - so, a kind of theropod with a long body, almost nonexistent arms, and thick, powerful legs. Rajasaurus in particular differed from other Abelisaurs in having particularly short legs, making it even more… sausage-like… in appearance than even its close relatives. It had a boxy head and thick neck, which would allow it to have a very powerful bite and strength in the neck to hold down prey. It had a strong sense of smell, as well, to help it to find prey from farther away - allowing it to set up an ambush for said prey when it got too close. It had horns on its forehead, made of bone from the nose, which was probably not extended by skin. It was also a lot shorter than other Abelisaurids - which means that it was only about 7 or so meters long, and maybe only two meters tall, if that. It really wouldn’t have stood much taller than an adult man. Rajasaurus had an especially short neck, which may have allowed it to grab onto prey even tighter than other Abelisaurids. It had very short, four-fingered hands, with claws on the first three of them. Though the legs of Rajasaurus are short, it did have very robust, thick toes, giving it more support on the ground. As an Abelisaurid, Rajasaurus was covered in scales all over its body, with potentially round bumpy bits of bone (osteoderms) interspersed among the scales.
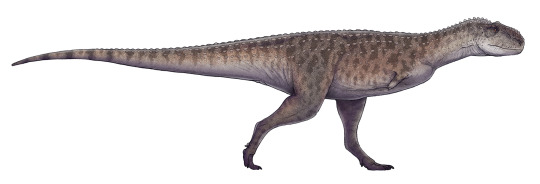
By Paleocolour, CC BY-SA 3.0
Diet: As a large Abelisaurid, Rajasaurus would have primarily fed upon larger herbivores, such as Titanosaurs.
Behavior: Abelisaurs were not the fastest animals - after all, if they ran too quicky, they wouldn’t have been very balanced - but what they were extremely agile turners. Being able to turn very quickly allowed them to be efficient ambush predators. Rajasaurus would have waited for prey to appear, and then charged - seemingly out of nowhere, by turning rapidly towards the prey - and then grabbing down onto the struggling prey with its strong, boxy jaws. Though Rajasaurus had small arms and hands, it traded those off for having a stronger neck - to better hold the prey steady with. Then, the stress would weaken the animal, along with blood loss. The horns on the head of Rajasaurus were probably for display and interaction between members of the species, with fights occurring to argue over carcasses or for mates via head-butting or neck-bashing. The horns would have also packed quite a bit of a cutting edge in these fights. It’s also possible that bright colors could have been used on the horns to display. While it doesn’t seem likely that Rajasaurus was particularly social, it did probably take care of its young, and may have formed small family groups while they grew up, in order to protect the young from the many other predators around.

By ДиБгд, CC BY-SA 3.0
Ecosystem: The Lameta Formation is a fascinating Late Cretaceous environment due to the fact that it was a grassland! Grasses evolved sometime in the Cretaceous period. While they did not spread rapidly until the Cenozoic, they did seem to be present in quite a few areas during the Cretaceous (to the point of some hadrosaur relatives evolving to eat them). India at the time was an isolated island, so grasses were able to thrive and diversify more there than in other locations where they did not gain as immediate of a foothold. There was a large amount of volcanic activity nearby, which probably added to its extensive biodiversity (before it made many of the animals present go extinct, via the explosion of the Deccan Traps). It was a lush environment filled with grasses resembling modern rice, flowers, algae, and ferns. This was an environment filled with many lakes, surrounded by extensive mud that lead to its fossil preservation. Rajasaurus was certainly not the only dinosaur of this environment, either! There were even other Abelisaurids - Indosaurus, Indosuchus, Rahiolisaurus, Lametasaurus, and their close relatives the (potentially piscivorous) Noasaurids such as Ornithomimoides, Laevisuchus, and Dryptosauroides. There were also other theropods, probably also Ceratosaurs - Jubbulpuria, Coeluroides, and Orthogoniosaurus. With all of these predators and fishermen, it makes sense that there were a lot of large herbivores for them to feed upon! And there seem to be at least three different kinds of titanosaurs there - Titanosaurus, a dubiously known one; Jainosaurus, a slightly better known one; and Isisaurus, the best known one that seems to be one of the main features of the environment. There was also potentially an ankylosaur - Brachypodosaurus! As for non-dinosaurs, there were large snakes, Sanajeh and Madtsoia; a variety of turtles; and some Dyrosaurid crocodylomorphs!

By José Carlos Cortés
Other: Rajasaurus was one of the Majungasaurines, which had longer holes in their snouts in front of their eyes than their close relatives the Carnotaurines - aka, they had lighter skulls - and having small crests widening the front of their heads; and, in general, longer snouts. This group of Abelisuarids underwent extensive island hopping, reaching places like India via rafting and other journeys across the ocean. Rajasaurus, being one of them, had ancestors that underwent such a journey!
~ By Meig Dickson
Sources Under the Cut
Brookfield, M. E.; Sanhi, A. (1987). "Palaeoenvironments of the Lameta beds (late Cretaceous) at Jabalpur, Madhya Pradesh, India: Soils and biotas of a semi-arid alluvial plain". Cretaceous Research. 8 (1): 1–14.
Carrano, M. T.; Sampson, S. D. (2008). "The phylogeny of Ceratosauria" (PDF). Journal of Systematic Palaeontology. 6 (2): 183–236.
Cau, A., F. M. Dalla Vecchia, and M. Fabbri. 2012. Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco. Acta Palaeontologica Polonica 57(3):661-665
Delcourt, R. (2018). "Ceratosaur Palaeobiology: New Insights on Evolution and Ecology of the Southern Rulers". Scientific Reports. 8 (9730).
Filippi, L. S., A. H. Méndez, R. D. Juárez Valieri and A. C. Garrido. 2016. A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids. Cretaceous Research 60:209-219
Furtado, M. R., C. R. A. Candeiro, and L. P. Bergqvist. 2013. Teeth of Abelisauridae and Carcharodontosauridae cf. (Theropoda, Dinosauria) from the Campanian- Maastrichtian Presidente Prudente Formation (southwestern São Paulo State, Brazil). Estudios Geológicos 69(1):105-114
Gianechini, F. A., S. Apestteguia, W. Landini, F. Finotti, R. J. Valieri and F. Zandonai. 2015. New abelisaurid remains from the Anacleto Formation (Upper Cretaceous), Patagonia, Argentina. Cretaceous Research 54:1-16
Grillo, O. N.; Delcourt, R. (2016). "Allometry and body length of abelisauroid theropods: Pycnonemosaurus nevesi is the new king". Cretaceous Research. 69: 71–89.
Kapur, V. V.; Khosla, A. (2016). "Late Cretaceous terrestrial biota from India with special reference to vertebrates and their implications for biogeographic connections". Cretaceous Period: Biotic Diversity and Biogeography. 71: 161–172.
Mohabey, D. M. 1989. The braincase of a dinosaur from the Late Cretaceous Lameta Formation, Kheda District, Gujarat, western India. Indian Journal of Earth Sciences 16(2):132-135
Mohabey, D. M. (1996). "Depositional environment of Lameta Formation (late Cretaceous) of Nand-Dongargaon inland basin, Maharashtra: the fossil and lithological evidences". Memoirs of the Geological Survey of India. 37: 1–36.
Mohabey, D. M.; Samant, B. (2013). "Deccan continental flood basalt eruption terminated Indian dinosaurs before the Cretaceous–Paleogene boundary" (PDF). Geological Society of India Special Publication (1): 260–267.
Novas, F. E., S. Chatterjee, D. K. Rudra and P. M. Datta. 2010. Rahiolisaurus gujaratensis, n. gen. n. sp., a new abelisaurid theropod from the Late Cretaceous of India. In S. Badyopadhyay (ed.), New Aspects of Mesozoic Biodiversity. Lecture Notes in Earth Sciences 132. Springer-Verlag, Berlin 45-62
Persons IV, W. S.; Currie, P. J. (2011). "Dinosaur speed demon: the caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids". PLoS One. 6 (10): e25763.
Prasad, V.; Strömberg, C.A.; Leaché, A.D.; Samant, B.; Patnaik, R.; Tang, L.; Mohabey, D.M.; Ge, S.; Sahni, A. (2011). "Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae". Nature Communications. 2: 480.
Prasad, V., C. A. E. Strömberg, H. Alimohammadian, A. Sahni. 2005. Dinosaur Coprolites and the Early Evolution of Grasses and Grazers Science 310 (5751): 1177-1180.
Prasad, G. V. R., and A. Sahni. 2009. Late Cretaceous continental vertebrate fossil record from India: palaeobiogeographical insights. Bulletin de la Société géologique de France 180(4):369-381
Ratsinbaholison, N. O., R. N. Felice, and P. M. O'Connor. 2016. Ontogenetic changes in the craniomandibular skeleton of the abelisaurid dinosaur Majungasaurus crenatissimus from the Late Cretaceous of Madagascar. Acta Palaeontologica Polonica 61(2):281-292
Rogers, Raymond R.; Krause, David W.; Curry Rogers, Kristina; Rasoamiaramanana, Armand H.; Rahantarisoa, Lydia. (2007). "Paleoenvironment and Paleoecology of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar". In Sampson, S. D.; Krause, D. W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology. Society of Vertebrate Paleontology Memoir 8. 27. pp. 21–31.
Sahni, A. 1972. Paleoecology of Lameta Formation at Jabalpur (M.P.). Current Science 41(17):652
Sahni, A. 1984. Cretaceous-Paleocene terrestrial faunas of India: lack of endemism during drifting of the Indian Plate. Science 226(4673):441-443
Sampson, Scott D.; Witmer, L. M. (2007). "Craniofacial anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar". In Sampson, S. D.; Krause, D. W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar (PDF). Journal of Vertebrate Paleontology. Society of Vertebrate Paleontology Memoir 8. 27. pp. 32–102.
Sereno, P. C.; Wilson, J. A.; Conrad, J. L. (2004). "New dinosaurs link southern landmasses in the Mid–Cretaceous". Proceedings of the Royal Society B. 271 (1546): 1325–1330.
Sereno, P. C.; Brusatt, S. L. (2008). "Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger" (PDF). Acta Palaeontologica Polonica. 53 (1): 15–46.
Sonkusare, H.; Samant, B.; Mohabey, D. M. (2017). "Microflora from Sauropod Coprolites and Associated Sedimentsof Late Cretaceous (Maastrichtian) Lameta Formation of Nand-Dongargaon Basin, Maharashtra". Geological Society of India. 89 (4): 391–397
Tandon, S. K.; Sood, A.; Andrews, J. E.; Dennis, P. F. (1995). "Palaeoenvironments of the dinosaur-bearing Lameta Beds (Maastrichtian), Narmada Valley, Central India". Palaeogeography, Palaeoclimatology, Palaeoecology. 117 (3–4): 153–184.
Tortosa, T., E. Buffetaut, N. Vialle, Y. Dutour, E. Turini and G. Cheylan. 2013. A new abelisaurid dinosaur from the Late Cretaceous of southern France: palaeobiogeographical implications. Annales de Paléontologie.
Vianey-Liaud, M.; Khosla, A.; Garcia, G. (2003). "Relationships between European and Indian dinosaur eggs and eggshells of the oofamily Megaloolithidae". Journal of Vertebrate Paleontology. 23 (3): 575–585.
Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; Noto, Christopher N. (2004). "Dinosaur distribution". In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606.
Wilson, J. A., P. C. Sereno, S. Srivastava, D. K. Bhatt, A. Khosla and A. Sahni. 2003. A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India. Contributions from the Museum of Paleontology, University of Michigan 31(1):1-42
Wilson, J. A.; Mohabey, D. M.; Peters, S. E.; Head, J. J. (2010). "Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India". PLoS One. 8 (3): e1000322.
#Rajasaurus narmadensis#Rajasaurus#Abelisaurid#Dinosaur#Ceratosaur#Palaeoblr#Factfile#Theropod Thursday#Carnivore#India & Madagascar#Cretaceous#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
191 notes
·
View notes
Photo

Island Weirdness #04 -- Gargantuavis philoinos
By far the biggest island in the Late Cretaceous European archipelago, the Ibero-Armorican island (sometimes also known as the Ibero-Occitan island) was made up of most of the Iberian Peninsula and France and was larger than modern-day Madagascar.
Around 73-71 million years ago one of the residents of this island was the aptly-named Gargantuavis -- the largest known Mesozoic bird, and probably an example of island gigantism.
Although only known from a few isolated bones, it’s estimated to have been slightly larger than a modern cassowary, somewhere in the region of 2m tall (6′6″). At that size it would have also been secondarily flightless, which is surprising for a bird that was living alongside larger fast-moving theropods like abelisaurs.
Not much else is known about it due to the scarce remains, but it seems to have had a long slender neck and probably had a small head. Its hips were fairly broad, suggesting it wasn’t capable of running very fast, and it was likely a slow-moving herbivore that was a fairly rare member of its ecosystem.
Exactly where it belongs in the bird evolutionary tree is also unclear, with the best current guess being “some sort of euornithean”.

#island weirdness 2019#science illustration#paleon#paleoart#palaeoblr#gargantuavis#euornithes#avialae#bird#stem-bird#dinosaur#flightless#art#ibero-armorcian island#island weirdness part 1
149 notes
·
View notes
Text
The Science of Jurassic Park & More: Part Three
In today’s article, we will be continuing the final part of our deep-dive into the world of palaeontology with Doctor David Button from London’s Natural History Museum.
In Part Two, David talked about his favourite dinosaurs, whether we are still discovering new dinosaurs, and much more. If you missed it, click here to take a read.
Many of the images in the article ahead are also courtesy of our friends at Jurassic Vault – so go show them some love if you haven’t already!
Without any further ado, let’s kick off our final part to this series by talking about the accuracy of the fan favourite Spinosaurus!
Image from Jurassic Park Wiki.
How accurate was the Spinosaurus we saw in Jurassic Park 3 (2001) at the time of release?
The Spinosaurus of Jurassic Park III was actually reasonably accurate at the time of release, as it was not well known at the time. Its main problem was the orientation of the hands – as with most of the other theropods in the franchise, it was depicted with permanently pronated “bunny hands”, even though theropods were unable to pronate their hands into such a position.
How do you see/research intelligence of extinct animals that died millions of years ago?
We can get an idea of the cognition of an extinct animal by taking a cast of the interior surfaces of the braincase – producing what is known as an endocast. This used to be performed by injecting latex into a dinosaur braincase and then cutting it open. Fortunately, however, these days we are able to produce these digitally by CT-scanning dinosaur braincases. The endocast gives us a replica of the shape of the brain and surround soft tissues.
From this, we can examine the shape of the brain and its constituent lobes to get an idea of the sensory abilities of an animal, and from the dimensions of the inner ear we can even calculate the range of sounds an animal could hear. The overall size and shape of the brain also give us a rough idea of the animal’s intelligence relative to that of other creatures. We can then also compare this with other evidence of dinosaur behaviours, such as associations of skeletons and footprints, to get an idea of their cognition and behavioural complexity. However, it should be stated that brain size does not necessarily correlate tightly with intelligence, and certainly many animals with small brains are still capable of complex cognition and behaviours. Consequently, we can only get a vague impression of just how smart dinosaurs may have been.
Would a Mosasaurus have slept in the same way a Whale sleeps?
Sleeping poses a problem for marine animals that need to come up for air, but cannot haul out onto land. Whales have solved this by only putting part of the brain to sleep at a time, still allowing them to surface. Sea turtles, however, have different solutions. Sometimes they sleep floating at the surface. Otherwise, however, sea turtles are able to exploit their low metabolisms. They slow their metabolic rate and oxygen use so as to not require another breath for hours, allowing them to sleep for that time under the surface.
It is difficult to say for sure Mosasaurs would have slept. However, there is evidence that Mosasaurs were warm-blooded, with a relatively high metabolic rate. Consequently, I think they would have been more likely to sleep like a whale than like a turtle.
Stiggy.....is it a growth phase of the pachycephalosaur?
Stygimoloch, Dacrorex and Pachycephalosaurus are differently-sized genera of pachycephalosaur, all known from the same time and place. Work on the bone histology of the animals has shown that the known specimen of Dacrorex is an infant, Stygimoloch a juvenile, and Pachycephalosaurus an adult. This suggests that they may all be growth phases in the life of a single animal. However, it may equally mean that they are three different animals that just happen to be known from different life stages. Unfortunately, pachycephalosaur remains are generally rare, and with such small sample sizes it is difficult to satisfactorily prove this problem one way or the other.
However, that being said, it seems more likely to me that these dinosaurs are all different life stages of the same animal than that there were three very similar pachycephalosaurs living in the same time and place. Hence, I think it is best to assume this option, until more specimens let us test this more thoroughly. I do admit that this is disappointing, though, as Stygimoloch was one of my favourite dinosaurs.
Is it true that we don’t know what Carnotaurus’ feet look like? And it’s tail?
It is true that specimens of Carnotaurus that we do have do not preserve these parts of the skeleton, so we do not know exactly what they looked like. However, Carnotaurus was an Abelisaurid, and these parts of the skeleton are known from other Abelisaurs, such as Majungasaurus, allowing us to fill them in with a reasonable degree of accuracy. This suggests that the tail of Carnotaurus would have been rather unremarkable. The hind legs would have been quite stocky, but what we do have from Carnotaurus indicates that its legs would have been relatively longer than those of Majungasaurus, and so Carnotaurus would have been a more competent pursuit predator.
The question I always wanted to ask is. How come they decided to go for reptiles instead of birds for their Jurassic Park films and why are the raptors are bigger than their real-life counterpart?
Jurassic Park originally used reptiles as a model for dinosaurs partially as this was the prevailing view when the book was first written, and also as that was the public expectation of dinosaurs. Nonetheless, the book and first movie movie did draw heavily on birds for some of the depicted behaviours, so that each were quite radical when they came out. Since then, however, the Jurassic Park franchise has stuck with more reptilian dinosaurs despite the science leaving that far behind. This is probably partially to preserve continuity, and partially because the public still, unfortunately, expects and prefers their dinosaurs to be as reptilian as possible when watching movies.
Velociraptor was originally scaled-up to more resemble the size of Deinonychus by Michael Crichton when he was writing Jurassic Park. This partially followed the unusual classification of Dromaeosaurids by the palaeontologist Greg Paul, who considered Velociraptor mongoliensis and Deinonychus antirrhopus to both belong to the same genus. Jurassic Park followed this, featuring Deinonychus, but calling it Velociraptor Antirrhopus. This also explains why the featured Velociraptor skeleton was discovered by Grant in the USA, as opposed to Mongolia, where Velociraptor actually lived. However, no other palaeontologists agree with this classification, not the least because the two animals lived on separate continents, 30 million years apart! Nonetheless, Crichton decided to stick with the name Velociraptor, because he considered it more dramatic.
When it came to adapting Jurassic Park for film, the raptors were increased in size again. This was due partially to Spielberg, reportedly, being unimpressed with the size of Deinonychus. However, it was also due to the discovery of Utahraptor at around the same time – it had not yet been described and named, but there were reports of large raptor bones being found at the time that inspired the production crew of the movie.
If an eccentric millionaire was able to bring back dinosaurs and make a Jurassic Park, what is the probability of them breaking out and creating all the chaos seen in the films?
That’s a big if! However, if they did, I am sure that dinosaurs would break out every so often. How much chaos were then able to cause would be directly down to how well-prepared the millionaire had been. If they had stringent systems in place, an escaped dinosaur should be quickly containable – although their large size and strength would pose significant challenges, they would not be any more vicious or intelligent than other zoo animals. However, if they had under prepared with inadequate staff, and over reliance on untested automated systems and untrustworthy employees, then, well…
Considering how obvious this is, and I've asked this before, does the don't move thing for Tyrannosaurus sound ridiculous and is 12.5 meters the plausible maximum for the king of the Cretaceous? If a Rex went 32 mph, would it trip over?
You are correct in that vision-based eyesight for a large predator like Tyrannosaurus makes no sense at all. I am glad the series has been downplaying this since Jurassic Park.
How quickly Tyrannosaurus could run remains a strong point of contention within palaeontology. Still, most estimates do hover around the region of a running speed of 11-13 metres per second (~25 mph) – this seems sensible to me. At higher speeds the problem is not that Tyrannosaurus would necessarily fall over, but that if it did happen to trip, the head would hit the ground with such momentum that it would be fatal. Hence, it is possible that it could have briefly used such a burst of speed when closing in on prey, but it would not have used such speeds during normal locomotion or pursuit.
Are Giganotosaurus and Carcharodontosaurus considered Allosaurid or Allosauroid? And if suddenly someone brings dinosaurs to life through their original DNA, would you use frog or bird DNA, to make them as accurate as possible? And is the Jurassic Park T-Rex a bit shoehorned and shrinkwrapped?
Carcharodontosaurids such as Giganotosaurus and Carcharodontosaurus itself are Allosauroids. However, they are not also Allosaurids, but are instead more closely related to Neovenatorids: these two groups together form the Carcharodontosauria.
Were it possible to extract DNA from dinosaur fossils to revive them, I would certainly use bird DNA to plug the gaps. This would both make the resulting dinosaurs more accurate and also, due to using a more closely related animal, make it more likely that the created embryos would be viable in the first place. Unfortunately, though, it does not appear possible for DNA to survive for that long in fossils.
‘Shrink-wrapping’ is a term used to describe palaeoart that does not put enough flesh and other soft tissues on a dinosaur’s bones. As you say, the Tyrannosaurus in Jurassic Park has suffered a little bit of shrinkwrapping – the spine, shoulder blade, ribs and hips are very obvious, as are some of the edges of the skull. However, overall, most of the dinosaurs in Jurassic Park are not too bad for shrinkwrapping – the Tyrannosaurus, for example, just looks underfed – this may be as the need to create a moving 3D model means the animators will get some kind of feel for the volumes of muscle that should be present. Still, most of the dinosaurs should have lips, and the Pterosaurs in Jurassic World were shrinkwrapped to a horrifying degree, as were the heads of the Apatosaurs in the same movie.
And which parts of the Jurassic Park T-Rex would you rather fix to give nod to accuracy?
I would draw upon evidence of the integument of tyrannosaurs to make it both more accurate, but also more visually interesting. Although we know that Tyrannosaurus was mostly covered in scales, we also know that it had some regions of naked skin (and, possibly, some feathers). Furthermore, we know that Tyrannosaurus’ face was armoured by large, overlapping scales – it would be nice to include these. Despite this, I do also think that Theropods such as Tyrannosaurus would have had lips, and so would include those. The ridges on Tyrannosaurus’ head were probably for display, and it would have had good colour vision, so brightly colouring those seems both plausible and visually interesting.
In addition, drawing on the above discussion about ‘shrinkwrapping’, I would beef Tyrannosaurus up a bit, especially around the legs and tail (both related to the leg muscles, but also likely fat stores in the tail as seen in modern reptiles). I tend to like my dinosaurs to be bulky, as we tend to estimate volumes of soft tissue in animals in general from their skeletons.
How much can you rely on fused bone sutures to understand a dinosaur’s growth stage? For example, why does a juvenile T. rex have fused sutures but appears to be young when looking at its histology?
The degree of sutural fusion in a dinosaur is often used to get a rough idea of life stage although, as you say, there are many caveats. In tyrannosaurs, for example, sutures in the axial and appendicular regions of the skeleton fused very early during growth, whereas many of those in the skull never fused, even in adulthood. The fusion of sutures between bones is governed, in part, on the forces acting upon them. Consequently, the stresses place on the tyrannosaur’s back and limbs could cause them to fuse early. By contrast, the open sutures between tyrannosaur facial bones served to help absorb and redistribute shocks encountered during feeding.
With those caveats in mind, bone histology remains the best tool to understand whether a dinosaur had reached adulthood. Still, unfused sutures in certain bones are still useful in showing us whether a dinosaur was still a juvenile, even if their fusion may not necessarily indicate that they were yet an adult. For example, the unfused scapulacoracoid of the mounted skeleton of Giraffatitan means that we know it was still growing, and was not fully adult.
Even though modern evidence suggests that adult tyrannosaurs probably had no feathers, does evidence suggest that young ones may have had feathers — possibly to help regulate heat?
We know that ancestral Tyrannosauroids – such as Dilong and Yutyrannus had feathers, and so they were secondarily lost in Tyrannosaurids, perhaps due to their large size obviating the need for insulation (which, indeed, would have caused heat stress in such large animals). Consequently, I think it is very plausible that tyrannosaur chicks would have had some kind of down-like coat to help keep them warm, especially in more northerly regions of tyrannosaur habitat. Indeed, it seems probable that the small, polar Tyrannosaurid Nanuqsaurus would have retained a feathery coat in adulthood, to help it last the cold winters.
It should also be noted that, although preserved skin impressions show that Tyrannosaurids such as Tyrannosaurus and Albertosaurus were predominately scaly, we still cannot rule out the presence of small regions of feathers, whether they were relicts of their evolutionary history or growth, or even for display.
What do we know of the lifespan of dinosaurs? Elephants have a gestational period of two years. Do we know anything about length of egg carrying or incubating time? Massive dinosaurs (Titanosaurs: how long until they reach adulthood? Would it be safe to assume they lived a long time?
We can get an idea of the age of a dinosaur specimen from looking at sections of its bones, especially the long bones of the limbs. In many cases, the bones of dinosaurs bones show growth rings – like those of trees – which can be counted to establish an age. Unfortunately, some dinosaurs – especially whilst young – grew too quickly to leave rings. In other cases, remodeling of bone structure after reaching adulthood also overpinted the rings. However, even then, the texture of bone indicates how rapidly the animal was growing when it died. Comparing these between differently-sized individuals of a dinosaur species allows us to establish growth curves. This, in turn, helps to establish when a dinosaur reached sexual maturity – as indicated by a slowdown in growth as resources are diverted for reproduction – and also the age at which maximum body size was achieved.
These results show that most dinosaurs grew fast – even large dinosaurs reached sexual maturity in their teens, and maximum body size in their twenties. Maximum lifespan is harder to establish, though, as it is impossible to verify whether any particular dinosaur died of old age. However, we know of some individuals that were at least 38 years old. However, it should be noted that very few dinosaurs would have reached old age, or even full body size – most would have died relatively young, due to their violent lifestyles. Once reaching full body size, however, they would be relatively safe from attack, and maximum lifespans of 50 years or more for large dinosaurs would seem likely.
Incubation times of dinosaur eggs can be calculated by counting growth rings in the teeth of dinosaur embryos. These indicate that incubation times in dinosaurs were variable. Most dinosaur eggs probably incubated for similar timescales to those seen in modern reptiles, taking from between 3-6 months to hatch. However, those of more birdlike dinosaurs, such as Troodon, hatched more quickly (but still more slowly than in modern birds), after only ~70 days.
Even though this means Titanosaur eggs may have taken several months of burial to hatch, the low investment a mother would have to put into each egg means that she would be able to lay multiple clutches per year, each containing as many as 30 eggs, for an annual total of perhaps as many as 200 eggs in the largest species. This is a much higher reproductive rate than a modern elephant, which has to invest 22 months of pregnancy, followed by 3-5 years of devoted parental care in a single calf. The lesser pressures of egg-laying relative to pregnancy are one reason why dinosaurs were able to reach much larger sizes than mammals such as elephants.
If the Baryonyx was around today would we see them in the swamps of Florida and or Louisiana?
Evidence from the isotopic composition of the bones of Spinosaurids such as Baryonyx, as well as from fish scales preserved in the animal’s stomach, indicate that it mostly ate fish and spent a large amount of its time in or around water. The warm, swampy environment of the Everglades would seem like ideal habitat for Baryonyx – and, indeed, is not too far from the wet environment of the Wealden Formation from which it is known. No Spinosaurids are currently known from North America, but that does not mean that Baryonyx would not be able to thrive were it somehow introduced there!
We really hope you found this three-piece series interesting! Make sure you also check out Part One and Part Two.
A massive thank you, again, to David for working on this series with us. We’ve found it incredibly interesting – and we hope you have too! Make sure to follow David on Twitter if you aren’t already, and stay tuned to The Jurassic Park Podcast for all the latest Jurassic Park news!
Written by: Tom Fishenden
#article#doctor david button#david button#ask a palaeontologist#ask a paleontologist#paleontologist#paleontology#science of jurassic park#tom fishenden#tom jurassic
0 notes
Link
Time for the next round!!! It’s time to vote in the NON-AVIAN THEROPOD GENUS that WILL be the competitor in this year’s Dinosaur March Madness!!!! All eligible genera ARE LISTED. Please READ the below information so that you make an informed voting choice! You have through January 14th!
HIGHLIGHTS & INELIGIBLES
Basal Theropods
Basal members of Theropoda that do not fit into more derived groups. Distinguishing characteristics of theropods include a wishbone. Theropods have remained persistently bipedal throughout their history. Ancestrally carnivorous, some theropods later adapted to omnivory or herbivory. Highlighted genera include Daemonosaurus (basal theropod with a short skull and protruding teeth), Liliensternus (medium-sized theropod, one of the largest known Triassic theropods from Europe and the best represented), Panguraptor(Coelophysoid known to not have had a head crest unlike other basal theropods), and Tawa (basal theropod that may represent the ancestral body plan of most other theropods). List of ineligible candidates: Dilophosaurus, Coelophysis
Misc. Tetanurans
Tetanuran theropods that don’t fit into more derived groups. Tetanurans are defined as all theropods more closely related to modern birds than to Ceratosaurus. Tetanurans possess stiffened tails and three-fingered hands. Highlighted genera include Australovenator (Megaraptoran with extremely flexible forelimbs), Megaraptor (Megaraptoran with large, sickle-shaped claws), Monolophosaurus (medium-sized theropod known from a nearly complete skeleton with a long crest on its skull), and Sinosaurus (theropod similar to Dilophosaurus known with an exceptionally well-preserved braincase). List of ineligible candidates: Cryolophosaurus
Basal Ceratosaurs
Theropods that are more closely related to Ceratosaurus than to modern birds. Ceratosaurs are known from the Early Jurassic to the Late Cretaceous. Ceratosaurs are known for elaborate crests and ridges on the skull, as well as reduced forelimbs, though the latter were more prominent in derived Ceratosaurs than in basal Ceratosaurs. Most were predators. Highlighted genera include Ceratosaurus (large ceratosaur with a prominent bladelike horn on the snout), Elaphrosaurus (slender ceratosaur with a notably shallow chest), Limusaurus (an herbivorous ceratosaur that gained a toothless beak in adulthood), and Masiakasaurus (small theropod with front teeth that projected outwards rather than vertically, probably used for fishing). List of ineligible candidates: None
Abelisauridae
Ceratosaurian theropod dinosaurs that first appeared in the Cretaceous. Towards the end of the Cretaceous, they became the main large predators in the Southern Hemisphere. Abelisaurid skulls tended to be tall in height and short in length, and often featured bony projections along the top. Abelisaurids possessed extremely short arms in proportion to their body size, with most lacking wrist bones. Highlighted genera include Abelisaurus (large abelisaurid known from a partial skull), Aucasaurus (medium-sized theropod with low brow ridges on its skull), Rugops (small abelisaur with a prominently ridged texture on top of the skull), and Rajasaurus (abelisaur with a single nose horn). List of ineligible candidates: Carnotaurus, Majungasaurus
Megalosaurs
All theropods more closely related to Megalosaurus than to modern birds. This group includes the Spinosaurs, large bodied theropods that had long crocodile-like snouts and tall projections along the spine forming a sail, and the megalosaurs, also large bodied theropods that had low, long skulls with almost no ornamentation. Highlighted genera include Suchomimus (large spinosaurid with long, slender jaws and a low sail on its back), Piatnitzkysaurus (medium-sized theropod with a preserved braincase), Torvosaurus (very large theropod with a kinked snout), and Megalosaurus (notable for being the first validly named non-avian dinosaur). List of ineligible candidates: Baryonyx, Spinosaurus
Carnosaurs
Predatory theropods classified as tetanurans more closely related to Allosaurusthan to modern birds. They ranged from the Jurassic to the Upper Late Cretaceous. Carnosaurs possessed distinctive characteristics such as large eyes with long, narrow skulls, as well as the thigh being longer than the shin. Highlighted genera include Acrocanthosaurus (very large theropod with tall projections on its spine), Neovenator (Allosauroid with complex passages for nerves in its snout that may have functioned as sensory organs), Yangchuanosaurus (large theropod with multiple hornlets and ridges on its skull), and Concavenator (had a tall, narrow pointed crest on its back and possible quill knobs on its forearms). List of ineligible candidates: Allosaurus, Carcharodontosaurus
Basal Coelurosaurs and Compsognathids
Mostly predatory theropods that are more closely related to modern birds than to Carnosaurs. A large number of feathered Coelurosaur specimens have been found, and it has been suggested that all Coelurosaurs possessed some stage of feathers. Notable characteristics include slender hands and feet as well as larger brains than other theropods. Compsognathids were small Coelurosaurs. They varied in general form, but both preserved scales and feathers are known for the group. Highlighted genera include Ornitholestes (small coelurosaurian with a retractable foot claw), Sinocalliopteryx (large Compsognathid with preserved protofeathers and gut contents), Zuolong (potentially the most basal coelurosaur known), Scipionyx (compsognathid with preserved soft tissue and internal organs), and Sciurumimus (coelurosaur with long fluffy feathers on its tail). List of ineligible candidates: Sinosauropteryx, Compsognathus
Tyrannosauroids
Coelurosaurian theropods that first appeared in the Jurassic and grew to be the dominant predators in the Northern Hemisphere by the end of the Cretaceous. Early Tyrannosauroids were small and possessed three-fingered hands while the later, more derived Tyrannosaurids grew massive with proportionally small arms, two-fingered hands, and bone-crunching jaws. Protofeathers have been found in at least two species, Yutyrannus and Dilong. Highlighted genera include Dilong (small tyrannosauroid known from nearly complete specimens, one preserved with protofeathers), Albertosaurus (large tyrannosauroid with skin impressions), Guanlong (small tyrannosauroid with a large crest on its skull), Daspletosaurus (large Tyrannosauroid with the longest forelimbs of any tyrannosaurid relative to body size), and Alioramus (medium-sized, long-snouted Tyrannosauroid known from scant remains). List of ineligible candidates: Tyrannosaurus, Yutyrannus
Ornithomimosaurs
Theropods that superficially resembled modern ostriches. They appeared in the Early Cretaceous and lasted into the Late Cretaceous. They possessed small skulls with large eye sockets and many later ornithomimosaurs had toothless beaks. Ornithomimosaurs also had long arms with strong claws, and are believed to have been omnivores or herbivores. Feathers have been found on several specimens, including some that have very ostrich-like plumage. Most ornithomimosaurs appear to have been built for fast running. Highlighted genera include Pelecanimimus (small ornithomimosaur with numerous small teeth), Dromiceiomimus (medium-sized ornithomimosaur known from many well-preserved specimens, including some with feathers), Rativates (small ornithomimosaur discovered in the 1930s but unrecognized as a new species until 2016), and Garudimimus (small ornithomimosaur with short heavy legs). List of ineligible candidates: Deinocheirus, Gallimimus
Therizinosaurs and Alvarezsaurs
Therizinosaurs were theropods from the Early to Late Cretaceous. They were herbivores that possessed massive claws on their hands. They also had modified hips that resembled those of ornithischians, robust hindlimbs, and protofeathers, as well as potentially more complex feathers. Alvarezsaurs were small, long-legged theropods. Later forms bore small, but strong forelimbs that seem to have been adapted for digging or tearing, with a claw being present only on the enlarged thumb. Alvarezsaurs had long, tube-like jaws with many teeth, which has been suggested to indicate that they fed on termites and other colonial insects. At least one specimen has preserved downy feathers. Highlighted genera include Beipiaosaurus (basal Therizinosaur preserved with several types of feathers), Falcarius (one of the basalmost therizinosaurs known, potentially more omnivorous than later forms), Segnosaurus (large Therizinosaurid with flattened claws), Haplocheirus (large basal Alvarezsauroid that still possessed three functional fingers), and Xixianykus (small Alvarezsaurid with extremely long legs and feet for its body size). List of ineligible candidates: Therizinosaurus, Shuvuuia, Mononykus, Nothronychus
Oviraptorosaurs
Maniraptoran theropod dinosaurs from Cretaceous Asia and North America. Derived forms are notable for having distinct beaked, parrot-like skulls that often bear some sort of crest. Oviraptorosaurs had short tails, and are known to have brooded over their nests of eggs. Feathers have been found in several forms, including attachment points for wing feathers along the forearm. Highlighted genera include Caudipteryx (small Oviraptorosaur preserved with complex wing and tail feathers), Avimimus (small Oviraptorosaur with fused hand bones and long legs, known from numerous specimens preserved as a bonebed), Khaan (Oviraptorid that might have had sexual dimorphism and known from complete remains), and Incisivosaurus (Oviraptorosaur with flattened, rodent-like front teeth). List of ineligible candidates: Citipati, Anzu
Basal Paravians and Troodontids
Basal members of Paraves that don’t fit into other groups. Most possessed a large claw on the second toe of each foot that was usually held off the ground when walking. Paravians generally have winged forelimbs, though the size of the wing relative to the body varies. Troodontids were small, birdlike theropods. They possessed skulls with densely-packed teeth and small sickle claws on their second toes, though these were not as large as the ones seen in dromaeosaurs. They are suggested to have had strong senses, most notably sight and hearing. Highlighted genera include Jianianhualong (small troodontid that preserves a feathered tail frond and bird-like wings), Mei (small troodontid known from two specimens preserved in sleeping posture), Zanabazar (the largest known Asian troodontid), and Scansoriopteryx (small paravian that had an extremely long third finger possibly supporting a membranous wing). List of ineligible candidates: Troodon, Yi
Dromaeosaurs
Small- to medium-sized theropods that possessed pennaceous feathers. Their most notable characteristic is the large, sickle claw on the second toe of each foot. This claw is theorized to have been used to help hold down prey when hunting. Many specimens have been found with complex feathers. Highlighted genera include Zhenyuanlong (medium-sized dromaeosaur preserved with massive wing feathers like birds), Halszkaraptor (small dromaeosaur that shows adaptations for semi-aquatic life like a goose), Buitreraptor (small dromaeosaur with an elongated snout suited to fishing), and Achillobator (very large dromaeosaur with rather stocky legs). List of ineligible candidates: Deinonychus, Utahraptor, Microraptor, Velociraptor, Dakotaraptor
Basal Avialans
Avialae includes modern birds and all dinosaur s more closely related to them than to “classic” dinosaurs such as Velociraptor and Troodon. Basal avialans were mostly small theropods similar to dromaeosaurids and troodontids. Some forms such as the Confuciusornithids and Sapeornis had very short tails, similar to modern birds. The ability to fly also became more widespread in Avialae, though these basal members were probably still relatively poor fliers compared to most modern flying birds. Highlighted genera include Jeholornis (relatively large, seed-eating avialan with unusual tail plumage), Serikornis (small avialan with “silky” feathers similar to those of Silkie chickens), Anchiornis (small avialan known from numerous well-preserved specimens, allowing for detailed reconstruction of its plumage and coloration), and Sapeornis (relatively large, seed-eating avialan that was probably adapted to soaring). List of ineligible candidates: Archaeopteryx, Balaur, Confuciusornis
Enantiornitheans
One of the most successful groups of Mesozoic avialans. Enantiornitheans were mostly small, tree-dwelling dinosaurs representing a broad diversity of ecologies. Most Enantiornitheans likely had flight abilities approaching those of modern birds, but they typically differed from modern birds in several notable ways, such as having teeth instead of beaks, as well as a relatively slow growth rate. Highlighted genera include Shanweiniao (enantiornithine with several pairs of elongate tail feathers, likely for display), Iberomesornis (one of the smallest known Mesozoic dinosaurs), Pengornis (relatively large enantiornithine with blunt teeth), Bohaiornis (relatively large enantiornithine that may have been raptorial), and Longipteryx (long-snouted enantiornithine). List of ineligible candidates: Avisaurus
Euornitheans
Euornithes includes modern birds and everything more closely related to them than to Enantiornitheans. Euornitheans differ from other Avialans in being able to spread and fold the fan of feathers on their tail, which helps them maneuver in flight. They are also beaked on at least their jaw tips, though many Mesozoic forms retained teeth further back in the jaws. The following are Euornitheans that don’t belong to the more specific group Neornithes (the least inclusive group including all modern birds). Highlighted genera include Ichthyornis (one of the first-described Mesozoic Euornitheans), Apsaravis (desert-dwelling Euornithean known from a nearly complete specimen), Patagopteryx (flightless Euornithean with stumpy feet), and Yanornis (fish-eating Euornithean known from many well-preserved specimens). List of ineligible candidates: Hesperornis
By @quetzalcoatlusnorthropi, @tacosaurusbell, @albertonykus, and @raptorcivilization
160 notes
·
View notes