#dromomeron
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
12.7% of mobile users access Tumblr.
Text

My last paleoart of 2024, here comes Coelophysis bauri and Dromomeron romeri from Ghost ranch.
47 notes
·
View notes
Text


Lagerpetidae
... is a family of basalavemetatarsalians. Though traditionally considered the earliest-diverging dinosauromorphs (reptiles closer to dinosaurs than to pterosaurs), fossils described in 2020 suggest that lagerpetids may instead be pterosauromorphs (closer to pterosaurs). Lagerpetid fossils are known from the Late Triassic of Argentina, Arizona, Brazil, Madagascar, New Mexico, and Texas. They were typically small, although some lagerpetids, like Dromomeron gigas and a specimen from the Santa Rosa Formation attributed to Dromomeron sp., were able to get quite large (femoral length 150–220 mm (5.9–8.7 in)). Lagerpetid fossils are rare; the most common finds are bones of the hindlimbs, which possessed a number of unique features...
Read more: Lagerpetidae - Wikipedia
illustrations: Nobu Tamura and Maurissauro
8 notes
·
View notes
Text
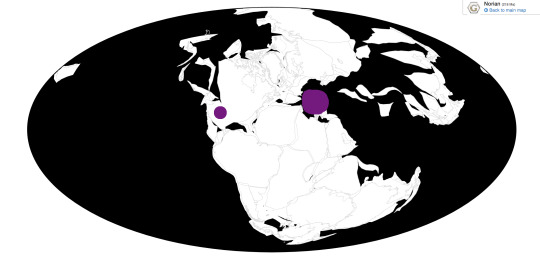
Dromomeron romeri, D. gregorii, D. gigas

By Jack Wood on @thewoodparable
PLEASE SUPPORT US ON PATREON. EACH and EVERY DONATION helps to keep this blog running! Any amount, even ONE DOLLAR is APPRECIATED! IF YOU ENJOY THIS CONTENT, please CONSIDER DONATING!
Name: Dromomeron romeri, D. gregorii, D. gigas
Name Meaning: Running Femur
First Described: 2007
Described By: Irmis et al.
Classification: Avemetatarsalia, Ornithodira, Dinosauromorpha, Lagerpetidae
Dromomeron is a genus of Dinosauromorph, specifically a Lagerpetid, a group of early-derived Dinosauromorphs from the Late Triassic. This group contains a handful of very small animals which were beginning to adapt for fast, upright, active movement - a foreshadowing of their later dinosaurian relatives. Dromomeron, specifically, lived about 220 to 211 million years ago, in the Norian age of the Late Triassic. It was about 1 meter long, and has three species assigned to it. D. romeri is from the Chinle Formation of New Mexico, known from a partial skeleton and other remains of hindlimb bones. D. gregorii hails from the Colorado City Formation of Texas, known from limb bones, and it was slightly older than D. romeri. D. gigas was found in the Quebrada del Barro Formation of Argentina. It, like other Ornithodirans, was an active animal, probably covered in filamentous integument, and would have lived alongside other Dinosauromorphs, including Dinosaurs proper, in complex Triassic ecological communities.
Source:
https://en.wikipedia.org/wiki/Dromomeron
Shout out goes to @cahethel1979!
#dromomeron#dinosauromorph#lagerpetid#dromomeron romeri#dromomeron gregorii#dromomeron gigas#palaeoblr#cahethel1979#dinosaur#paleontology#prehistory#prehistoric life#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#factfile#Dìneasar#דינוזאור#डायनासोर#ديناصور#ডাইনোসর#risaeðla#ڈایناسور#deinosor#恐龍
241 notes
·
View notes
Text
Vancleavea campi

By Tas Dixon
Etymology: Named after Phillip Van Cleave
First Described By: Long & Murry, 1995
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Proterochampsia?, Doswelliidae?
Status: Extinct
Time and Place: 228 to 203.6 million years ago, in the Norian of the Late Triassic

Vancleavea is best known from the Petrified Forest in Arizona and Ghost Ranch in New Mexico, but is found throughout the Chinle Formation in Arizona, New Mexico, Texas and Utah in the southern United States. It is also known from the Tecovas Formation in Texas, and the Redonda and Bull Canyon Formations in New Mexico.
Physical Description: Vancleavea is a peculiar reptile, even by Triassic archosauromorph standards. It effectively resembles an armoured, scaly eel with tiny legs, a long neck, the tail of a crocodile and a frighteningly fang-toothed skull. The largest complete specimen is approximately 1.2 metres (3.9 feet) long, although isolated pieces from other specimens indicate it could grow larger. Most of its body is covered in an array of overlapping bony osteoderms that come in 5 different shapes on the main body, throat, underbelly, limbs and tail (as well as possibly some strange spike-like osteoderms of unknown position). Uniquely, the osteoderms on the tail are tall and flat from side to side, forming a tall fin along the top of the tail for swimming. No other tetrapod—except for the closely related Litorosuchus—has a tail fin like this (usually they are made of tall neural spines from the tail bones).
Vancleavea’s skull is also unusual, as it is very robust and solid, despite the relatively thin and pointed snout. The back of the skull is boxy and has completely closed over the upper temporal fenestra, leaving just the very large lower temporal fenestra. The antorbital fenestra in front of the eyes, a typical feature of archosauriform reptiles, has also been lost, and the snout is short with nostrils that point directly upwards. The jaw bones of Vancleavea are heavily pitted and have a rough texture, which combined with the large teeth imply Vancleavea lacked lips and had exposed fangs, similar to crocodiles. Vancleavea had three fangs on each side of its jaws, one at the front, one on the lower jaw, and another on the upper jaw behind the bottom fang. The dentary fang slotted into a gap in the upper jaw when the mouth was closed, again similar to crocodiles.
The body of Vancleavea is long and tubular, enhanced by both the unusually long neck and the small size of its limbs. It’s unlikely Vancleavea’s limbs were strong enough to lift its body off the ground on land, and in fact the forelimb is so reduced that the shape of some its finger bones suggests it had even lost the claws on its hands. All in all Vancleavea is very clearly built to be semi-, if not almost entirely, aquatic, swimming with a sinuous, eel-like motion and rarely crawling onto land, perhaps only to lay its eggs.
Diet: The massive, serrated fangs and conical teeth indicate Vancleavea was carnivorous, likely feeding on fish and other aquatic prey, such as small temnospondyl amphibians.
Behavior: Vancleavea is almost certainly semi-aquatic, using its deep, finned tail to scull through the water similar to living crocodilians. It’s speculated that Vancleavea lived as an ambush predator, lunging forward to snag prey in its fangs. It lived in slow-moving freshwater environments, and its elongated, sinuous body would have been perfect for winding through plants and logs and other obstacles in the water.
Ecosystem: Vancleavea is known from much of the Chinle Formation, one of the most well known Late Triassic formations in the world, and its longevity means that it coexisted with a wide variety of other animals. There were of course the dinosaurs, including the likes of Coelophysis, Chindesaurus and Daemonosaurus, along with other dinosauriforms such as silesaurids and the lagerpetid Dromomeron, as well as early pterosaurs. Pseudosuchian archosaurs were incredibly abundant, such as the numerous herbivorous aetosaurs and Revueltosaurus, the dinosaur-like poposauroids including the herbivorous beaked Shuvosaurus and Effigia, as well as the predatory Poposaurus. Rauisuchid pseudosuchians were among the top predators, such as Postosuchus, while small long-legged crocodylomorphs like Hesperosuchus and Parrishia chased after smaller prey. Various other archosauromorphs were present too, including the bizarre drepanosaurs, herbivorous allokotosaurs such as Trilophosaurus and an unnamed azendohsaurid, and the strange armoured Doswellia, possibly Vancleavea’s closest relative. There was even an early turtle, Chinlechelys, one of the earliest known to have a fully formed shell, and a relative of the tuatara, Whitakersaurus. The only definitively known synapsid from the Chinle is the large herbivorous dicynodont Placerias.
In the water, Vancleavea would have coexisted with numerous species of giant phytosaurs—crocodile-like reptiles that may-or-may-not be pseudosuchians—such as Leptosuchus and Machaeoprosopus, as well as the predatory temnospondyl amphibian Anaschisma (formerly known as Koskinonodon). Vancleavea’s heavy armour may have been necessary protection against these predators, which could have had jaws almost as long as it was! Fish were abundant in the freshwater ecosystem, with various ray-finned fish such as the pike-like Saurichthys, the large lungfish Arganodus, freshwater sharks such as Xenacanthus, and even a freshwater coelacanth, Chinlea. Many of these fish grew to as large or larger than Vancleavea, and may have competed with it or even preyed upon it (at least as juveniles).
Vancleavea also coexisted with several other, more mysterious reptiles that are mostly only known from teeth. One of these, Acallosuchus, was discovered close by to the original specimen of Vancleavea. The skull and skeleton were originally suggested to belong to the same animal, but we now know Vancleavea had a very different skull to Acallosuchus. Supposedly its skull was long, pointed and triangular, with a knobbly texture on the bones. I say “supposedly” because the skull disintegrated while it was excavated, and all that’s left is a crude drawing and a few pieces of bone. The identity of Acallosuchus remains a mystery. Other mysterious species that Vancleavea coexisted with are the former ornithischians Tecovasaurus and Crosbysaurus, Colognathus, Palacrodon, the possibly venomous Uatchitodon, and the extremely enigmatic Kraterokheirodon known only from two teeth so unlike any known tetrapod alive or extinct it can only be classified as Amniota incertae sedis! Vancleavea was once in the same boat as all these creatures, known only by pieces of the skeleton and its armour, and with any luck, Acallosuchus and the other mystery creatures will end up like Vancleavea and turn up some complete skeletons one day.
Other: Vancleavea is known from throughout the Chinle Formation and several other localities in the southern US, however, most of these specimens consist of isolated bones and osteoderms. The Chinle Formation covers many tens of millions of years from top to bottom, and it’s unlikely the single species V. campi was that long-lived (its range spans a whole 20 million years!). Variations between some specimens suggest that there may be more diversity within Vancleavea then we currently recognise. However, until better specimens are found and can be compared, we can’t be sure how Vancleavea evolved and changed throughout the course of the Chinle.
Vancleavea has a fairly complicated history, the holotype was first discovered in the Petrified Forest by fossil collector Phillip Van Cleave, who lent it his name, but it wasn’t described and named until 1995 and still only consisted of a fragmentary skeleton. Nearly complete specimens from Ghost Ranch were discovered in 1982, but they wouldn’t be recognised as Vancleavea until decades later. In 2008, new specimens from the Petrified Forest were published, but Vancleavea still had to wait another year before the Ghost Ranch specimens were extensively described and its strangeness could be fully appreciated.
Until recently, Vancleavea was the only known stem-archosaur of its kind known in the world, and its relationship to other archosauriforms was enigmatic. However, the discovery of the closely related Litorosuchus from China indicates that the Vancleavea-family (“vancleaveans”) was indeed more diverse and widespread. Unlike Vancleavea, Litorosuchus was marine, and had a much longer, almost spinosaur-like snout. The position of Vancleavea in the archosauromorph tree is still uncertain, but it has been suggested to be a type of proterochampsian, a group of typically crocodile-like reptiles closely related to archosaurs proper. Particularly, it has been allied to the family Doswelliidae, a peculiar family of proterochampsians with similarly strange armour and skeletons. Regardless of where it truly belongs, Vancleavea is at least generally agreed to be a fairly derived archosauriform, quite close to the crown group of Archosauria, albeit having evolved down a very unusual, unique route all of its own.
~ By Scott Reid
Sources under the cut
Hunt, Adrian P.; Lucas, Spencer G.; Spielmann, Justin A. (January 2005). "The holotype specimen of Vancleavea campi from Petrified Forest National Park, Arizona, with notes on the taxonomy and distribution of the taxon". New Mexico Museum of Natural History and Science Bulletin. 29: 59–65.
Li, C.; Wu, X.-C.; Zhao, L.-J.; Nesbitt, S.J.; Stocker, M.R.; Wang, L.-T. (2016). "A new armored archosauriform (Diapsida: Archosauromorpha) from the marine Middle Triassic of China, with implications for the diverse life styles of archosauriforms prior to the diversification of Archosauria". The Science of Nature. 103 (95): 95.
Long, R. A. & Murry, P. A. (1995). "Late Triassic (Carnian and Norian) tetrapods from the southwestern United States". New Mexico Museum of Natural History and Science Bulletin. 4: 1–254.
Miller, Zachery (November 17 20016). D&D Monsters of the Triassic. Waxing Paleontologist.
Nesbitt, S. J.; Stocker, M. R.; Small, B. J.; & A. Downs (November 26, 2009). "The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes)". Zoological Journal of the Linnean Society. 157 (4): 814–864.
Parker, W. G.; Barton, B. (2008). "New information on the Upper Triassic archosauriform Vancleavea campi based on new material from the Chinle Formation of Arizona". Palaeontologia Electronica. 11 (3)
#vancleavea#archosauromorph#diapsid#reptile#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
287 notes
·
View notes
Text
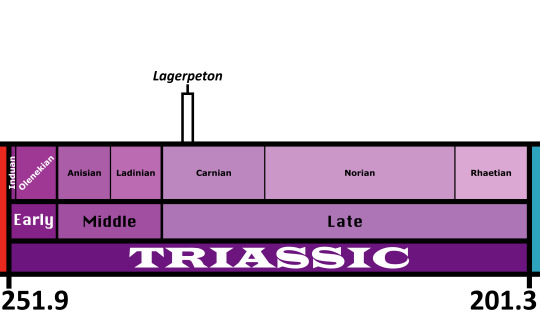
Lagerpeton

By Tas
Etymology: Rabbit Reptile
First Described By: Romer, 1971
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Crurotarsi, Archosauria, Avemetarsalia, Ornithodira, Dinosauromorpha, Lagerpetidae
Referred Species: L. chanarensis
Status: Extinct
Time and Place: About 235 to 234 million years ago, in the Carnian of the Late Triassic
Lagerpeton is known from the Chañares Formation in La Rioja, Argentina

Physical Description: Lagerpeton was named as the Rabbit Reptile, and for good reason - in a lot of ways, it represents a decent attempt by reptiles in trying to do the whole hoppy-hop thing. You might think that it resembles Scleromochlus in that way, and you’d be right! Scleromochlus and Lagerpeton are close cousins, but one is on the line towards Pterosaurs - Scleromochlus - and the other is on the line towards dinosaurs - Lagerpeton. So, hopping around was an early feature that all Ornithodirans (Dinosaurs, Pterosaurs, and those closest to them) shared. Lagerpeton itself was about 70 centimeters in length, with most of that length represented as tail; it was slender and lithe, built for moving quickly through its environment. It had a small head, a long neck, and a thin body. While it had long legs, it also had somewhat long arms, and while it may have been able to walk on all fours it also would have been able to walk on two legs alone. It was digitigrade, walking only on its toes, making it an even faster animal. Its back was angled to help it in hopping and running through its environment, and its small pelvis gave it more force during hip extension while jumping. In addition to all of this, it basically only really rested its weight on two toes - giving it even more hopping ability! As a small early bird-line reptile, it would have been covered in primitive feathers all over its body (protofeathers), though what form they took we do not know.
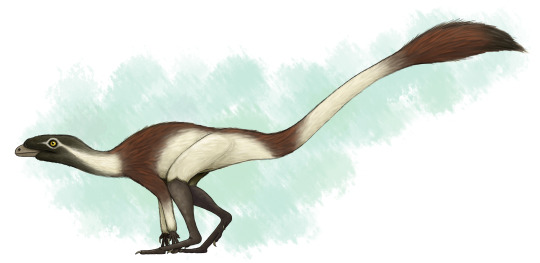
By Scott Reid
Diet: As an early dinosaur relative, it’s more likely than not that Lagerpeton was an omnivore, though this is uncertain as its head and teeth are not known at this time.
Behavior: Lagerpeton would have been a very skittish animal, being so small in an environment of so many kinds of animals - and as such, that hopping and fast movement ability would have aided it in escaping and moving around its environment, avoiding predators and reaching new sources of food (and, potentially, chasing after smaller food itself). Lagerpeton may have also been somewhat social, moving in small groups, potentially families, to escape the predators and chase after prey together, given its common nature in its environment. As an archosaur, Lagerpeton was more likely than not to take care of its young, though we don’t know how or to what extent. The feathers it had would have been primarily thermoregulatory, and as such, they would have helped it maintain a constant body temperature - making it a very active, lithe animal.
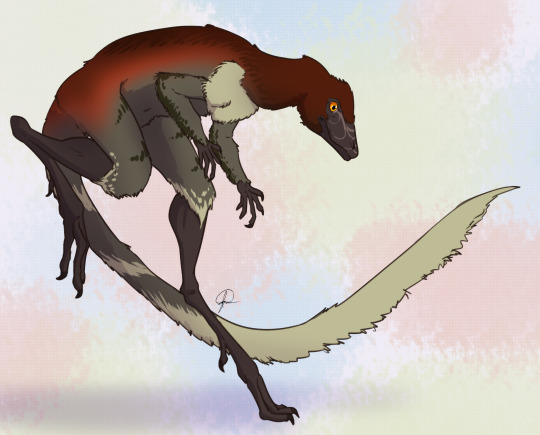
By José Carlos Cortés
Ecosystem: Lagerpeton lived in the Chañares environment, a diverse and fascinating environment coming right after the transition from the Middle to Late Triassic epochs. Given that the first true dinosaurs are probably from the start of the Late Triassic, this makes it a hotbed for understanding the environments that the earliest dinosaurs evolved in. Since Lagerpeton is a close dinosaur relative, this helps contextualize its place within its evolutionary history. This environment was a floodplain, filled with lakes that would regularly flood depending on the season. There were many seed ferns, ferns, conifers, and horsetails. Many different animals lived here with Lagerpeton, including other Dinosauromorphs like the Silesaurid Lewisuchus/Pseudolagosuchus and the Dinosauriform Marasuchus/Lagosuchus. There were crocodilian relatives as well, such as the early suchian Gracilisuchus and the Rauisuchid Luperosuchus. There were also quite a few Proterochampsids, such as Tarjadia, Tropidosuchus, Gualosuchus, and Chanaresuchus. Synapsids also put in a good show, with the Dicynodonts Jachaleria and Dinodontosaurus, as well as Cynodonts like Probainognathus and Chiniquodon, and the herbivorous Massetognathus. Luperosuchus would have definitely been a predator Lagerpeton would have wanted to get away from - fast!

By Ripley Cook
Other: Lagerpeton is one of our earliest derived Dinosauromorphs, showing some of the earliest distinctions the dinosaur-line had compared to other archosaurs. Lagerpeton was already digitigrade - an important feature of Dinosaurs - as shown by its tracks, called Prorotodactylus. These tracks also showcase that dinosaur relatives were around as early as the Early Triassic - and that their evolution, and the rapid diversification of archosauromorphs in general, was a direct result of the end-Permian extinction.
~ By Meig Dickson
Sources Under the Cut
Arcucci, A. 1986. New materials and reinterpretation of Lagerpeton chanarensis Romer (Thecodontia, Lagerpetonidae nov.) from the Middle Triassic of La Rioja, Argentina. Ameghiniana 23 (3-4): 233-242.
Arcucci, A. B. 1987. Un nuevo Lagosuchidae (Thecodontia-Pseudosuchia) de la fauna de los Chañares (Edad Reptil Chañarense, Triasico Medio), La Rioja, Argentina. Ameghiniana 24: 89 - 94.
Arcucci, A., C. A. Mariscano. 1999. A distinctive new archosaur from the Middle Triassic (Los Chañares Formation) of Argentina. Journal of Vertebrate Paleontology 19: 228 - 232.
Bittencourt, J. S., A. B. Arcucci, C. A. Marsicano, M. C. Langer. 2014. Osteology of the Middle Triassic archosaur Lewisuchus admixtus Romer (Chañares Formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs. Journal of Systematic Palaeontology: 1 - 31.
Brusatte, S. L., G. Niedzwiedzki, R. J. Butler. 2011. Footprints pull origin and diversification of dinosaur stem lineage deep into Early Triassic. Proceedings of the Royal Society B: Biological Sciences 278 (1708): 1107 - 1113.
Ezcurra, M. D. 2006. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28(4):649-684.
Ezcurra, M. D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriformes. PeerJ 4: e1778.
Fechner, R. 2009. Morphofunctional evolution of the pelvic girdle and hindlimb of DInosauromorpha on the lineage to Sauropoda (Thesis). Ludwigs Maximillians Universita.
Fiorelli, L. E., S. Rocher, A. G. Martinelli, M. D. Ezcurra, E. Martin Hechenleitner, M. Ezpeleta. 2018. Tetrapod burrows from the Middle-Upper Triassic Chañares Formation (La Rioja, Argentina) and its palaeoecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 496: 85 - 102.
Kammerer, C. F., S. J. Nesbitt, N. H. Shubin. 2012. The first Silesaurid Dinosauriform from the Late Triassic of Morocco. Acta Palaeontological Polonica 57 (2): 277.
Kent, D. V., P. S. Malnis, C. E. Colombi, A. A. Alcober, R. N. Martinez. 2014. Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proceedings of the National Academy of Sciences 111: 7958 - 7963.
Irmis, R. B., S. J. Nesbitt, K. Padian, N. D. Smith, A. H. Turner, D. T. Woody, and A. Downs. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317:358-361.
Jenkins, F. A. 1970. The Chañares (Argentina) Triassic reptile fauna. VII. The postcranial skeleton of the traversodontid Massetognathus pascuali (Therapsida, Cyondontia). Breviora 352: 1 - 28.
Langer, M. C., S. J. Nesbitt, J. S. Bittencourt, R. B. Irmis. 2013. Non-dinosaurian Dinosauromorphs. Geological Society London, Special Publications. 379 (1): 157 - 186.
Marsh, A. D. 2018. A new record of Dromomeron romeri Irmis et al., 2007 (Lagerpetidae) from the Chinle Formation of Arizona, U.S.A. PaleoBios 35:1-8.
Marsicano, C. A., R. B. Irmis, A. C. Mancuso, R. Mundil, F. Chemale. 2016. The precise temporal calibration of dinosaur origins. Proceedings of the National Academy of Sciences of the United States of AMerica 113 (3): 509 - 513.
Martz, J. W., and B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7:e7551:1-71.
Nesbitt, S. J., R. B. Irmis, W. G. Parker, N. D. Smith, A. H. Turner and T. Rowe. 2009. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. Journal of Vertebrate Paleontology 29(2):498-516.
Nesbitt, S. J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 353:1-292.
Perez Loinaze, V. S., E. I. Vera, L. E. Fiorelli, J. B. Desojo. 2018. Palaeobotany and palynology of coprolites from the Late Triassic Chañares Formation of Argentina: implications for vegetation provinces and the diet of dicynodonts. Palaeogeography, Palaeoclimatology, Palaeoecology 502: 31 - 51.
Rogers, R. R., A. B. Arcucci, F. Abdala, P. C. Sereno, C. A. Forster, C. L. May. 2001. Paleoenvironment and taphonomy of the Chañares Formation tetrapod assemblage (Middle Triassic), northwestern Argentina: spectacular preservation in volcanogenic concretions. Palaios 16: 461 - 481.
Romer, A. S. 1966. The Chañares (Argentina) Triassic reptile fauna. I. Introduction. Breviora 247: 1 - 14.
Romer, A. S. 1966. The Chañares (Argentina) Triassic reptile fauna. II. Sketch of the geology of the Rio-Chañares-Rio Gualo Region. Breviora 252: 1 - 20.
Romer, A. S. 1967. The Chañares (Argentina) Triassic reptile fauna. III. Two new gomphodonts, Massetognathus pascuali and M. teruggii. Breviora 264: 1 - 25.
Romer, A. S. 1968. The Chañares (Argentina) Triassic reptile fauna. IV. The dicynodont fauna. Breviora 295: 1 - 25.
Romer, A. S. 1969. The Chañares (Argentina) Triassic reptile fauna. V. A new chiniquodontid cynodont, Probelesodon lewisi - cynodont ancestry. Breviora 333: 1 - 24.
Romer, A. S. 1970. The Chañares (Argentina) Triassic reptile fauna. VI. A chiniquodont cynodont with an incipient squamosal-dentary jaw articulation. Breviora 344: 1 - 18.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. VIII. A fragmentary skull of a large thecodont, Luperosuchus fractus. Breviora 373: 1 - 8.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. IX: The Chanares Formation. Breviora 377: 1 - 8.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. X. Two new but incompletely known long-limbed pseudosuchians. Breviora 378:1-10.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. XI. Two new long-snouted thecodonts, Chanaresuchus and Gualosuchus. Breviora 379: 1 - 22.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XII. The post cranial skeleton of the thecodont Chanaresuchus. Breviora 385: 1 - 21.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XIII. A fragmentary skull of a large thecodont, Luperosuchus fractus. Breviora 389: 1 - 8.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. Lewisuchus admixtus, gen. et sp. Nov., a further thecodont from the Chañares beds. Breviora 390: 1 - 13.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XV. Further remains of the thecodonts Lagerpeton and Lagosuchus. Breviora 394: 1 - 7.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XVI. Thecodont classification. Breviora 395:1-24.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XVII. The Chañares gomphodonts. Breviora 396: 1 - 9.
Romer, A. S. 1973. The Chañares (Argentina) Triassic reptile fauna. XVIII. Probelesodon minor, a new species of carnivorous cynodont; family Probainognathidae nov. Breviora 401: 1 - 4.
Romer, A. S., and A. D. Lewis. 1973. The Chañares (Argentina) Triassic reptile fauna. XIX. Postcranial materials of the cynodonts Probelesodon and Probainognathus. Breviora 407: 1 - 26.
Romer, A. S. 1973. The Chañares (Argentina) Triassic reptile fauna. XX. Summary. Breviora 413: 1 - 20.
Sereno, P. C., and A. B. Arcucci. 1994. Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. Journal of Vertebrate Paleontology 14(1):53-73.
#Lagerpeton#Dinosaurmorph#Lagerpetid#Triassic#Palaeoblr#Triassic Madness#Ornithodiran#Triassic March Madness#South America#Omnivore#Prehistoric Life#Prehistory#Palaeontology#Lagerpeton chanarensis#dinosaur#paleontology#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#factfile
365 notes
·
View notes
Text
Drepanosaurus unguicaudatus

By Scott Reid
Etymology: Sickle Reptile
First Described By: Pinna 1979
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Protorosauria?, Drepanosauromorpha, Elyurosauria, Drepanosauridae, Megalancosaurinae
Time and Place: 212 to 205 million years ago, from the Norian to the Rhaetian ages of the Late Triassic

Drepanosaurus is known from the Zorzino Limestone and Dolomia di Forni Formations of Italy, and the Siltstone and Petrified Forest Members of the Chinle Formation in New Mexico
Physical Description: Drepanosaurus is the Titular Member of the Drepanosaurs, aka the “Monkey Reptiles”! These weirdos got their name for a reason - they were extremely well adapted for climbing trees, to the point of having prehensile tails like many monkeys. Drepanosaurus had a long, slender body, with a very long and thick tail for grabbing onto branches and moving around the forest. It had a long, thin neck and a small skull on the end of it, which was superficially similar to the skull of a bird - it was round on the back end, with huge eyes and a small pointed snout. Still, that head looked more like the head of a chick any number of nonavian dinosaurs, rather than like birds. It had long, robust limbs, with claws on the digits. Most notably, one front claw on each Drepanosaurus hand was huge, used for aiding in gripping branches more easily. It had an arched shoulder region, and in general very high spines on its vertebrae. This made it look quite odd indeed - a lumpy, lengthy reptile that sort of looks like what would happen if you tried to combine a woodpecker, a monitor lizard, a monkey, and a bison into one small reptilian creature that lived in the trees of the Triassic. It was weird, okay. It was just weird. The tail may have even ended in a claw - that’s right, a claw - to help it grab onto branches better. And all of this weirdness was packed into a body only about 45 to 50 centimeters long - a truly specialized representative of Triassic Oddities.
Diet: Drepanosaurus was most likely an insectivore, using the claws to help get out various insects from tree bark.
Behavior: Drepanosaurus was a weirdo, but a weirdo with a purpose. All of these weird traits came together in one wild animal to aid it in climbing among trees, gathering food, and moving nimbly in the forest. The small, pointed mouth aided in digging out food from crevices and burrows, which it could open up bigger using the large claws. Its long and skinny neck also helped it to reach spots where insects were hiding, rather than wasting energy to move its entire body to where food was on the tree. The large, splayed out feet and strong claws were able to wrap around tree trunks, gripping the wood tightly and helping Drepanosaurus stay stabilized. The prehensile tail helped Drepanosaurus go from branch to branch and move among the tree canopy, and the claw would help to keep the tail in place during particularly slippery or dangerous swings.
Ecosystem: Drepanosaurus lived in a variety of environments, most importantly the Triassic shores of Northern Italy, around tidal basins and seasonal lagoons. In the Zorzino beach formation, it shared its home with its close cousins, Megalancosaurus and Vallesaurus; as well as the early Pterosaurs Bergamodactylus, Peteinosaurus, and Eudimorphodon; the very long-snouted Phytosaur Mystriosuchus; the marine reptile Endennasaurus; the Placodont Psephoderma; the Aetosaur Aetosaurus; the Tuatara Diphydontosaurus; the Tanystropheid Langobardisaurus; and plenty of fish such as Legnonotus, Gibbodon, Sargodon, Pholidorhynchodon, Parapholidophorus, Pholidoctenus, Dapedium, Dandya, and the shark Pseudodalatias. This was a very active and diverse fauna, placing Drepanosaurus in one of the truly uniquely Triassic locations of the end of the Period.
Dolomia di Forni was no sneeze either, a Tidal Basin surrounded by coniferous trees. Here we have some preservation of things that Drepanosaurus was likely to eat, such as the spider Friularachne. There were also plenty of crustaceans, including decapods and crab like things that would have been crawling all over the tidal pools. Fish were very common too, such as the Coelacanth Holophagus, the sharp fish man Saurichthys, other Ray-Finned fish such as Eopholidophorus and Sargodon, the shark Pseudoalatias, and the Conodont Epigondolella. There was also the Tanystropheid Langobardisaurus again, as well as the other Drepanosaur Megalancosaurus. However, the true heroes of the environment were the early pterosaurs - the whole gang was there, including Preondactylus, Eudimorphodon, Austriadactylus, and Seazzadactylus. It was just a huge chaos of flying reptiles, which Drepanosaurus would have had a tough time avoiding!
Drepanosaurus has also been found in the Siltstone and Petrified Forest environments of the Chinle Formation in North America. Now, even in the context of Pangea this was a long distance away, but it’s possible they migrated there during the end of the Triassic period. These were thick coniferous forest environments with a severely dry and severely wet season, indicating that Drepanosaurus really adapted to a new cliimate, and quickly. Here there were famous animals such as the dinosaurs Coelophysis, Daemonosaurus, Chindesaurus, Tawa, the Silesaurid Eucoelophysis, the Lagerpetid Dromomeron, Pseudosuchians like Hesperosuchus, Effigia, Vivaron, and the Aetosaur Typothorax; the swimming Archosauromorph Vancleavea, Tuataras like Whitakersaurus, Phytosaurs like Redondasaurus and Rioarribasuchus, and even other Drepanosaurs like Avicranium (which might actually be what all the specimens of Drepanosaurus in the Chinle are, so take this with a grain of salt). This is a very diverse ecosystem that has been touched on elsewhere on WTF Triassic, so I will leave it at that!
Other: Oh ye Drepanosaurs. With heads so birdlike and bodies so weird. Much like other triassic weirdos, BANDits (“scientists” who insist, though literally all evidence points to the contrary, that birds could not have evolved from dinosaurs), you have been fodder for speculation for ages. However, you have almost nothing in common with birds - even the skull is only similar on the outside, and lacks many of the features seen in bird skulls (that are, however, found in dinosaur skulls). In fact, many of these creatures were grouped together in a group called Avicephala, which was eventually abandoned as it was found that most of them were not closely related. That said, it was then found that Drepanosaurs were Archosauromorphs, showing that weird arboreal lifestyles were a trend in the Ruling Reptile group, rather than a unique feature of the birds. Though, it is possible that Drepanosaurs are actually very early Diapsids (reptiles proper), and thus may have evolved as early as the Permian - and somehow survived into the Triassic. More fossils are needed, but for now, they remain a fascinating and unique Triassic group, highlighting the weirdness of this timeperiod in a way few other animals can.
~ By Meig Dickson
Sources Under the Cut
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution 1-698.
Colbert, E. H. 1947. The little dinosaurs of Ghost Ranch. Natural History 59(9):392-399-427-428.
Dalla Vecchia, F. M. 1991. Notes on the stratigraphy, sedimentology and paleontology of the Dolomia di Forni (Upper Triassic) of the Rio Seazza Valley (Preone, Friuli-Venezia Giulia)]. Gortania - Proceedings of the Friulian Museum of Natural History 12 (90) : 7-30.
Irmis, R. B., S. J. Nesbitt, K. Padian, N. D. Smith, A. H. Turner, D. T. Woody, and A. Downs. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317:358-361.
Harris, J. D., A. Downs. 2002. A drepanosaurid pectoral girdle from the Ghost Ranch (Whitaker) Coelophysis Quarry (Chinle Group, Rock Point Formation, Rhaetian), New Mexico. Journal of Vertebrate Paleontology 111: 247 - 264.
Pinna, G. 1979. Drepanosaurus unguicaudatus, nuovo genere e nuova specie di Lepidosauro del trias alpino. Atti della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 121: 181 - 192.
Pinna, G. 1986. On Drepanosaurus unguicaudatus, an Upper Triassic lepidosaurian from the Italian Alps. Journal of Paleontology 60 (5): 1127 - 1132.
Pinna, G. 1988. Un nuovo esemplare giovanile di Drepanosaurus unguicaudatus del Norico di Val Preone (Udine) [A new juvenile specimen of Drepanosaurus unguicaudatus from the Norian of the Preone Valley, Udine]. Atti della Società Italiana di Scienze Naturali - Museo civico di Storia Naturale di Milano 128:80-84.
Pinna, G., and S. Nosotti. 1989. Anatomia, morfologia funzionale e paleoecologia del rettile placodonte Psephoderma alpiunum Meyer, 1858. Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale de Milano 25(2):1-50.
Pritchard, A. C., S. J. Nesbitt. 2017. A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Open Science 4 (10): 170499.
Renesto, S. 1994. The shoulder girdle and anterior limb of Drepanosaurus unguicaudatus (Reptilia, Neodiapsida) from the Upper Triassic (Norian) of Northern Italy. Zoological Journal of the Linnean Society 111 (3): 247 - 264.
Renesto, S., J. A. Spielmann, S. G. Lucas, G. T. Spagnoli. 2010. The Taxonomy and paleobiology of the Late Triassic (Carnian-Norian: Adamnian-Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha). New Mexico Museum of Natural History and Science Bulletin 46: 1 - 81.
Senter, P. 2004. Phylogeny of Drepanosauridae (Reptilia: Diapsida). Journal of Systematic Palaeontology 2(3):257-268.
Stefani, M., P. Arduini, A. Garassino, G. Pinna, G. Teruzzi, G. L. Trombetta. 1991. Palaeoenvironment of extraordinary fossil biotas from the Upper Triassic of Italy. Atti della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 132 (24): 309 - 335.
#drepanosaurus#drepanosaur#diapsid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
283 notes
·
View notes
Text
Silesaurus opolensis

By José Carlos Cortés
Etymology: Silesian Reptile
First Described By: Dzik, 2003
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Silesauridae, Sulcimentisauria
Time and Place: Around 230 million years ago, in the Carnian of the Late Triassic
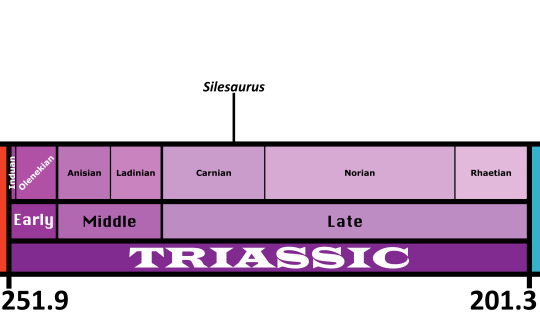
Silesaurus is known from the Drawno Beds Formation of Poland

Physical Description: Silesaurus was a very common almost-dinosaur - specifically, the titular member of the Silesaurids - from Poland in the middle of the Triassic period. It was a quadrupedal animal, about 2.3 meters long and small enough that it most likely would have been covered in fluff. Like other Silesaurids, it was long - with a long neck and a long tail, and long limbs easily reaching to the ground. Like other Silesaurids, it almost had its legs directly underneath its body - it didn’t seem to have an open acetabulum, but rather a partially open one, though Silesaurids are something of a confusing case on this regard. It had a small head, with a little beak in the front, and small sharp conical teeth. In addition, it had very large eyes, giving it distinctly good eyesight, as well as a decent sense of smell. As a quadruped, its front limbs were rather extended, while the hind limbs did a lot of the weight support.
Diet: Interestingly enough, though for a long time it was thought that Silesaurus was an herbivore, recent fossil poop studies have indicated it was an insectivore - similar to living birds.

By Ripley Cook
Behavior: If Silesaurus really was an insectivore, it would have spent much of its time moving around in its environment, examining it closely for insects and pecking at the ground with its small beak for food. This pecking may have even looked a lot like that of living birds - the large eyes probably would have prevented the eyes from moving much, like in living birds, and as such Silesaurus would have had to bob its head in order to see. It would have been very careful in its environment, making sure to look for food and avoid predatory dangers in its environment. It would probably have been social - corroborated by the fact that it was very common in its environment - and taken care of its young to some extent. As an Archosaur, it would have been warm blooded as well, and very active.
Ecosystem: Silesaurus lived in a Mediterranean-esque scrub environment, with summer monsoon seasons and dry winters. The swampy coniferous forest and fernlands it lived in had a wide variety of animals, placing Silesaurus in a unique ecosystem of the Carnian age. This is actually quite important, given that the Carnian Pluvial Event - a minor mass extinction that helped to spur Dinosaurian evolution - occurred around the same time, leading to Silesaurus and its contemporaries being a snapshot of this change. A variety of invertebrates lived alongside this animal, giving it plenty of food, as well as Temnospondyls such as Metoposaurus and Cyclotosaurus, the Aetosaur Stagonolepis, the Phytosaur Parasuchus, the Rauisuchid Polonosuchus, the Sharovpterygid Ozimek, and probably some Tuataras and maybe even a Pterosaur. A small community, but a beautiful portrait of this time period, with its fair share of weirdos and precursors. And Silesaurus would have had a lot to look out for with Polonosuchus at just the right size to attack it.
Other: Do not let Silesaurus’s weird similarities to birds fool you - it wasn’t an early bird, but a showcase of how bird-like traits kept popping up over and over again in the line of animals leading up to them. In fact, the evolution of such a specialized feeding system is notable in such an early member of the Ornithodirans. Silesaurids in general are extremely interesting because of their mixture of characters - similar to dinosaurs, but not quite there; with legs almost directly underneath the body, but other traits that match the later Ornithischians. The problem with Ornithischians - the bird-hipped dinosaurs - is that they have a bit of a ghost lineage, aka even though they should have showed up at the same time as Theropods and Sauropodomorphs, they don’t until the Early Jurassic. This isn’t as big of a gap as some other examples, but it is notable. Silesaurids have come out as Ornithischians just enough times in phylogenetic studies to bring it up as a question - are they almost-dinosaurs? The first bird-hips (without having bird-hips)? Something else entirely? More research is necessarily, but one thing is for sure - this group of weird dinosaurian experiments is one to keep an eye on, especially as we learn more and more about them.
~ By Meig Dickson
Sources Under the Cut
Cau, A. 2018. The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana 57 (1): 1 - 25.
Dzik, J. 2003. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology 23(3):556-574.
Dzik, J. and T. Sulej. 2007. A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Palaeontologia Polonica 64:3-27.
Ezcurra, M. D. 2006. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28(4):649-684.
Griffin, C. T., and S. J. Nesbitt. 2016. The femoral ontogeny and long bone histology of the Middle Triassic (?late Anisian) dinosauriform Asilisaurus kongwe and implications for the growth of early dinosaurs. Journal of Vertebrate Paleontology 36(3):e1111224:1-22.
Irmis, R. B., S. J. Nesbitt, K. Padian, N. D. Smith, A. H. Turner, D. T. Woody, and A. Downs. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317:358-361.
Jagt, J. W. M., G. Hebda, S. Mitrus, E. Jagt-Yazykov, A. Bodzioch, D. Konietzko-Meier, K. Kardynał, K. Gruntmejer. 2015. Field Guide, Conference Paper, European Association of Vertebrate Palaeontologists XIII Annual Meeting.
Kammerer, C. F., S. J. Nesbitt, and N. H. Shubin. 2012. The first basal dinosauriform (Silesauridae) from the Late Triassic of Morocco. Acta Palaeontologica Polonica 57(2):277-284.
Langer, M. C., M. D. Ezcurra, J. S. Bittencourt and F. E. Novas. 2010. The origin and early evolution of dinosaurs. Biological Reviews 85:55-110.
Martínez, R. N., and O. A. Alcober. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 4(2 (e4397)):1-12.
Martz, J. W., and B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7:e7551:1-71.
Nesbitt, S. J., C. A. Sidor, R. B. Irmis, K. D. Angielczyk, R. M. H. Smith and L. A. Tsuji. 2010. Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature 464:95-98.
Nesbitt, S. J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 353:1-292.
Qvarnström, M., J. V. Wernström, R. Piechowski, M. Tałanda, P. E. Ahiberg, G. Niedźwiedzki. 2019. Beetle-bearing coprolites possibly reveal the diet of a Late Triassic dinosauriform. Royal Society Open Science 6 (3): 181042.
#silesaurus#silesaurus opolensis#dinosauromorph#palaeoblr#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
244 notes
·
View notes
Text
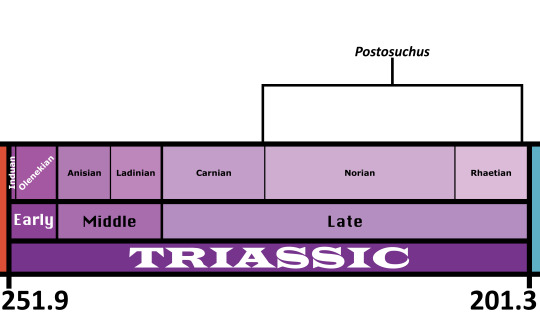
Postosuchus

By Ripley Cook
Etymology: Crocodile from Post, Texas
First Described By: Chatterjee, 1985
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Crurotarsi, Archosauria, Pseudosuchia, Suchia, Paracrocodylomorpha, Loricata, Rauisuchidae
Referred Species: P. kirkpatricki, P. alisonae
Status: Extinct
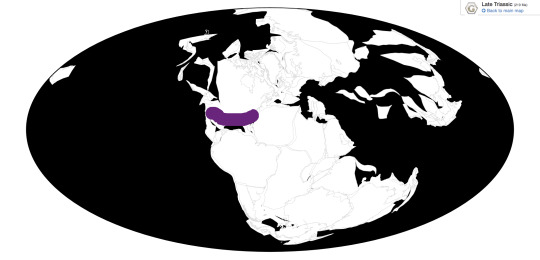
Time and Place: 227 to 201 million years ago, from the Norian to the Rhaetian of the Late Triassic.
Postosuchus is known from Arizona, New Mexico, Texas, and North Carolina.
Physical Description: Postosuchus was one the larger pseudosuchians of the Triassic, growing up to 6 meters (20 feet) in length. Its head was robust and rectangular in side view, with large spikelike teeth. The top of the skull has a flattened dorsal surface and roughened “brow ridges” that may have been covered with keratin in life. The body and tail were long and covered with osteoderms, which have prserved in the fossil record. Its forelimbs were much shorter than its hindlimbs and its hands were relatively small. Some of the fingers were slightly reduced in size, foreshadowing would happen to tyrannosaurs much later. The legs were longer, but shorter than those of theropod dinosaurs, giving Postosuchus a lower-slung stance. The hands and feet had five digits each.
Diet: Postosuchus was an apex predator. Its diet probably included dicynodonts, early dinosaurs, herbivorous pseudosuchians, and other archosauromorphs such as allokotosaurs.
Behavior: Based on how small the arms were relative to the hindlimbs, it is likely Postosuchus was bipedal, and probably even been an obligate biped based on the anatomy of its hips and limbs. It is not very well-built for pursuit, so Postosuchus was likely an ambush predator. It likely had well-developed senses of sight and smell. Like modern crocodilians, it may have been a largely solitary creature, with few Postosuchus in the area at a time. It is unknown whether or not it marked its territory with pee as in Walking with Dinosaurs.
Ecosystem: When Postosuchus lived, North America was arid, being inland on Pangaea. Most Postosuchus fossils come from the Chinle Formation of New Mexico and Arizona. Chinle was crawling with pseudosuchians: these include aetosaurs such as Typothorax and Desmatosuchus, the small herbivorous Revueltosaurus, the strangely coelurosaur-like Shuvosaurus, and the leggy “sphenosuchian” Hesperosuchus. Postosuchus also lived alongside early theropods like Coelophysis and Chindesaurus, the dinosauromorph Dromomeron, phytosaurs such as Smilosuchus, Leptosuchus and Machaeroprosopus, the bizarre archosauromorphs Trilophosaurus and Vancleavea, the long-necked tanystropheid Tanytrachelos, a drepanosaur, dicynodonts such as Placerias, cynodonts, lungfish, early sharks, and temnospondyls such as Koskinonodon and Apachesaurus. Postosuchus is also known from the coeval Dockum Group of Texas, which had many of the same fauna.
Meanwhile, in North Carolina, Postosuchus lived alongside other pseudosuchians such as Dromicosuchus and Stegomus, dicynodonts, the cynodont Plinthogomphodon, and temnospondyls. It would have been further inland, and thus even more arid.
Other: Sankar Chatterjee originally described Postosuchus in 1985 as the ancestor of tyrannosaurs. Unfortunately, Chatterjee is kind of an idiot. It was very soon after recognized to be an extinct relative of crocodilians. Chatterjee also referred several juvenile skeletons, which are now recognized as the adults of Shuvosaurus.
~ By Henry Thomas
Sources under the cut
Chatterjee, S. 1985. Postosuchus, a new thecodontian reptile from the Triassic of Texas and the origin of tyrannosaurs. Philosophical Transactiosn of the Royal Society of London B 309: 395-460.
Long, R.A., Murray, P.A. 1995. Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States. Bulletin of the New Mexico Museum of Natural History and Science 4: 1-254.
Weinbaum, J.C. 2011. The skull of Postosuchus kirkpatricki (Archosauria: Paracrocodyliformes) from the Upper Triassic of the United States. PaleoBios 30(1): 18-44.
Weinbaum, J.C. 2013. Postcranial skeleton of Postosuchus kirkpatricki (Archosauria: Paracrocodylomorpha), from the Upper Triassic of the United States. In: Nesbitt, S.J., Desojo, J.B., Irmis, R.B. eds. Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and their Kin. Geological Society, London, Special Publications 379: 525-553.
#postosuchus#rauisuchian#pseudosuchian#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
212 notes
·
View notes
Text
Xenacanthus

By @stolpergeist
Etymology: Weird spine
First Described By: Beyrich, 1848
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Chondrichthyes, Elasmobranchii, Euselachii, Selachimorpha, Xenacanthida, Xenacanthidae
Referred Species: X. atriossis, X. comrpessus, X. decheni, X. denticulatus, X. erectus, X. gibbosus, X. gracilis, X. howsei, X. indicus, X. laevissimus, X. latus, X. ludernensis, X. moorei, X. ossiani, X. ovalis, X. parallelus, X. parvidens, X. ragonhai, X. robustus, X. serratus, X. slaughteri, X. taylori
Status: Extinct
Time and Place: 295 to 208 million years ago, from the Sakmarian of the Early Permian to the Rhaetian of the Late Triassic.
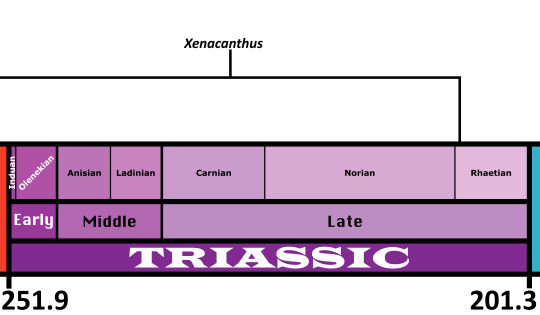
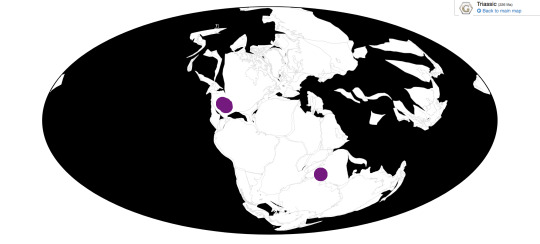
In the Triassic, Xenacanthus is known from New Mexico, Texas, and India.
Physical Description: Xenacanthus gets its name from the very large and narrow spike protruding out of the back of its head. Seriously, what’s up with that. Nobody really knows. Besidse that, the thing most people talk about are its teeth. Xenacanthid teeth are distinctively-shaped: they had a large oval base with two prongs shooting out, making the tooth look V-shaped. But beyond this, Xenacanthus is still interesting. The body is long and shallow, with ribbonlike dorsal and caudal fins that form an almost continuous fin going down the back and back up the bottom of the animal. The pectoral and pelvic fins are short and very broad.
Diet: Xenacanthus’s bizarre teeth may hint at a diet of crunchier foods. It likely ate fish and small invertebrates in the waters it lived, as well as potentially the carcasses of anything that drowned.
Behavior: Xenacanthus’s body is not well-equipped for fast swimming. Its pectoral and pelvic fins almost look limblike, and may have potentially been used to help maneuver (or “walk”) around river bottoms like modern epaulette sharks. Xenacanthus likely sat in the muddy bottoms of the rivers it lived in, waiting for prey to swim on by. Although it lived in rivers, Xenacanthus may have migrated to the sea to spawn like its relative Orthacanthus (and some living eels). The purpose of the spike on the back of the head is completely unknown, but it may have been used for display, defense, or stabilizing swimming.
Ecosystem: Xenacanthus lived in shallow freshwater environment such as seasonal rivers. It’s known from two areas in the Triassic: the southwestern United States, and India. In the southwestern US, it lived alongside the lungfish Ceratodus, various ray-finned fish, temnospondyls such as Metoposaurus and Koskinonodon, early dinosaurs like Coelophysis, dinosauromorphs such as Dromomeron, pseudosuchians such as Typothorax, Shuvosaurus, Postosuchus and Poposaurus, phytosaurs such as Rutiodon and Smilosuchus, and various other bizarre archosauromorphs like Vancleavea, Trilophosaurus, and Tanytrachelos. If this sounds familiar, this is because we’ve already featured lots of critters from Late Triassic North America. Lots of familiar faces.
Meanwhile in India, Xenacanthus lived alongside the hybodontid shark Polyacrodus, Ceratodus again, and the temnospondyl Buettneria. These rivers would have been frequented by thirsty amniotes such as the rhynchosaur Hyperodapedon, the protorosaur Malerisaurus, aetosaurs, the early dinosaur Alwalkeria, dicynodonts, the cynodont Exaeretodon, and that weirdo, you know it, you love it, Shringasaurus.
Other: Xenacanthus evolved in the Permian, and amazingly made it through the Great Dying! Sure, they were a lot more common in the Permian, but xenacanthids made it through to the end of the Triassic (when they were apparently done in by the end-Triassic extinction).
~ By Henry Thomas
Sources under the cut
Beck, K.G., Soler-Gijon, R., Carlucci, J.R., Willis, R.E. 2014. Morphology and Histology of Dorsal Spines of the Xenacanthid Shark Orthacanthus platypternus from the Lower Permian of Texas, USA: Palaeobiological and Palaeoenvironmental Implications. Acta Palaeontologica Polonica 61(1): 97-117.
Bhat, M.S. 2015. A new and diverse Late Triassic fish assemblage from India. International Conference on Current Perspectives and Emerging Issues in Gondwana Evolution, Lucknow.
Lucas, S.G., Heckert, A.B., Hotton, N. 2002. The rhynchosaur Hyperodapedon from the Upper Triassic of Wyoming and its global biochronological significance. New Mexico Museum of Natural History and Science Bulletin 21: 149-156.
Palmer, D. 1999. The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. Marshall Publishing, London.
Pauliv, V.E., Dias, E.V., Sedor, F.A., Ribeiro, A.M. 2014. A new Xenacanthiformes shark (Chondrichthyes, Elasmobranchii) from the Late Paleozoic Rio do Rasto Formation (Parana Basin), Southern Brazil. Anais da Academia Brasileira de Ciencias 86(1).
#xenacanthus#xenacanthid#shark#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
160 notes
·
View notes
Text
Triopticus primus

By Tyler Young/ @cry-olophosaurus
Etymology: Three eyes
First Described By: Stocker et al. 2016
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes incertae sedis
Referred Species: T. primus
Status: Extinct
Time and Place: 227 to 226 million years ago, in the Norian of the Late Triassic

Triopticus is known only from the Otis Chalk in Texas, the United States

Physical Description: Triopticus is a very puzzling and frustratingly enigmatic reptile, primarily because it is only known from the back portion of its skull and nothing else, leaving the shape of its body and even the size and shape of its snout a complete and utter mystery. We can’t even rely on phylogenetic bracketing to give us a guide of how it may have looked because its relationships cannot be pinned down beyond Archosauriformes. Even its body size is unclear, but it wasn’t a very big animal at least—the back of its skull could fit in the palm of your hand, so maybe around a metre long.
Nonetheless, the piece of skull that is known is very odd indeed. The back of its skull is remarkably similar to those of pachycephalosaurid dinosaurs, in that Triopticus was also a “bone head” with a thick bony “dome” over its head—it even flares out over the back and sides of the head like in pachycephalosaurs. Unlike pachycephalosaurs, however, the “dome” of Triopticus was not a single smooth structure, but was actually made up of five fused bosses on the bones of the skull that have clumped together to form a single bony dome-like structure. But, of course, the dome of Triopticus is even stranger than that of the pachycephalosaurs’. Right in the centre of the dome, between the bosses, there is a hole that drops right down to the roof of the skull. Like a doughnut. Why? No one knows. The hole happens to sit over right where the pineal gland could have been, and so the hole may have opened for a “third eye” on the top of its head, like many modern lizards and the tuatara have (this is where it gets its name!). And yet, the opening for the third eye was lost in the ancestor of Archosauriformes, so we shouldn’t expect Triopticus to have one in the first place. For that matter, the texture of the bone inside the hole is exactly the same as it is on the inside, implying it was covered in hard, opaque keratin like the rest of the dome. So Triopticus probably really did just have a hole in its head, you have to wonder how it kept that thing clean, or stop it from collecting rainwater.
As for the rest of the body, as frustratingly unclear as its relationships are, we can at least make an educated guess based on other archosauriforms. It was probably quadrupedal, with not very long sprawled to semi-erect legs, a long tail, and maybe some osteoderms running down its back. Of course, considering how weird just the back of its head is, and how other Triassic Weirdos have surprised us, I wouldn’t be surprised if Triopticus turned out to be weird all over. Maybe it was a biped with tiny forelimbs to really take the mickey out of the pachycephalosaur-convergance, who knows. (As it happens, the shape of its inner ear might suggest it really was bipedal, but the evidence is tenuous.)
Diet: Without any jaws and teeth, the diet of Triopticus is a mystery. All other known non-archosaur archosauriforms were carnivores, so it’s quite possible Triopticus was too. But there’s a first time for everything, and maybe the rest of its skull was just as abberant as its dome.
Behavior: Any and all speculations on the behaviour of Triopticus has to revolve around its dome. Fortunately, domes are interesting structures in animals, so there’s a decent amount to say. The obvious interpretation is that the dome convergently evolved for the same purposes suggested for pachycephalosaurs: head-butting. The utility of pachycephalosaur domes has been hotly debated, and presumably the same arguments apply here. Alternatively, the dome could have been just used for display. Either case implies Triopticus were social animals, either using the dome for visual communication or in confrontations between them. Triopticus had particularly big eyes and optic nerves for an archosauriform, so good vision must have been important for whatever it was doing. As before, any other behaviours are a mystery, although as an archosauriform we can reasonably speculate that it laid eggs and probably cared for its young to an extent.
Ecosystem: In the Otis Chalk, Triopticus co-existed with various other stem-archosaurs, including the herbivorous allokotosaur Trilophosaurus, the heavily armoured and boxy Doswellia, the phytosaurs Parasuchus and Angistorhinus, and three species of aetosaur. It also coexisted with at least four dinosaurmorphs, including the lagerpetid Dromomeron, a silesaurid, and the predatory theropods Chindesaurus and Lepidus. There was also the large predator Poposaurus, and even an ornithomimid-like shuvosaurid here too. This ecosystem in some ways was almost like a premonition of the later Cretaceous ecosystems, with phytosaur-like crocodiles, aetosaur-like ankylosaurs, poposaurid-like theropods, shuvosaurid-like ornithomimids, and even the Triopticus-like pachycephalosaurs. Whether Triopticus filled a similar ecological role to the pachycephalosaurs, and not just visual, is up for debate, and its lifestyle remains unknowable.
Other: The discovery of Triopticus and its uncanny similarity to pachycephalosaurs prompted a statistical analysis comparing the body types of various Triassic archosaurs and stem-archosaurs to those of Cretaceous dinosaurs. What they found was substantial overlap, so the eerie similarity between certain Triassic reptiles and dinosaurs wasn’t just people seeing things. The fact simply seems to be that Triassic stem- and crown-archosaurs had already evolved many of the distinctive body-types first known in—and thought to be characteristic of—dinosaurs. Triopticus now adds pachycephalosaurs into the “Triassic did it” roster, which at the time of discovery left only the giant sauropodomorphs, maniraptorans, and ceratopsians as the only ones not represented in the Triassic. Then Shringasaurus rolled around...
~ By Scott Reid
Sources under the cut
Georgi, J.A., Sipla, J.S., Forster, C.A. (2013). "Turning Semicircular Canal Function on Its Head: Dinosaurs and a Novel Vestibular Analysis". PLoS ONE. 8 (3): e58517.
Goodwin, M.B., Horner, J.R. (2004). "Cranial histology of pachycephalosaurs (Ornithischia: Marginocephalia) reveals transitory structures inconsistent with head-butting behavior". Paleobiology. 30 (2): 253–267.
Hieronymous, T.L., Witmer, L.M., Tanke, D.H., Currie, P.J. (2009). "The Facial Integument of Centrosaurine Ceratopsids: Morphological and Histological Correlates of Novel Skin Structures". Anatomical Record. 292 (9): 1370–1396.
Hullar, T.E. (2006). "Semicircular canal geometry, afferent sensitivity, and animal behavior". The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology. 288A (4): 466–472.
Nesbitt, S.J. (2011). "The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades". Bulletin of the American Museum of Natural History. 352: 1–292.
Stocker, M.R., Nesbitt, S.J., Criswell, K.E., Parker, W.G., Witmer, L.M., Rowe, T.B., Ridgely, R., Brown, M.A. (2016). "A Dome-Headed Stem Archosaur Exemplifies Convergence among Dinosaurs and Their Distant Relatives". Current Biology. 26 (19): 2674–2680.
Voogd, J., Wylie, D.R.W. (2004). "Functional and anatomical organization of floccular zones: A preserved feature in vertebrates". Journal of Comparative Neurology. 470 (2): 107–112.
#Triopticus#Triopticus primus#archosauriform#diapsid#archosauromorph#palaeoblr#triassic madness#triassic march madness#Triassic#reptile#prehistoric life#prehistory#paleontology
191 notes
·
View notes
Text
Effigia okeeffeae

By Stolpergeist
Etymology: Ghost
First Described By: Nesbitt and Norell, 2006
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Disapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Archosauria, Pseudosuchia, Suchia, Paracrocodylomorpha, Poposauroidea, Shuvosauridae
Referred Species: E. okeeffeae
Status: Extinct
Time and Place: Around 205 million years ago, in the Rhaetian of the Late Triassic
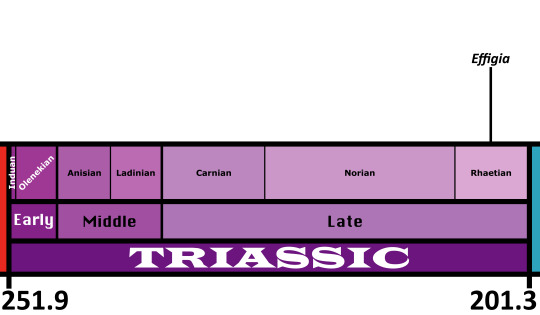
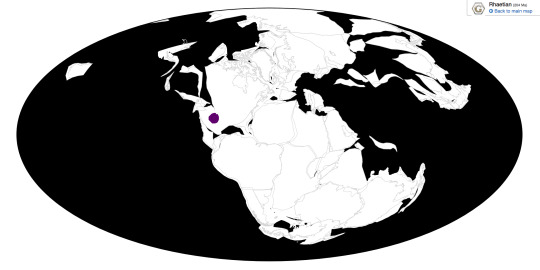
Effigia was found at Ghost Ranch in the Chinle Formation in New Mexico
Physical Description: Effigia was a small, bipedal animal that remarkably resembled dinosaurs it lived with at the time. In total, from head to tail, it would have been about two meters long. It walked on two legs that were held directly underneath the body, and it had small arms that weren’t used in supporting its weight. It had a long body, with a decently sized tail and long neck. The head of Effigia was small and narrow, ending in a beak and having no teeth whatsoever. So, in short, it looked like the later Limusaurus, except it wasn’t feathered - and it wasn’t a dinosaur! This is the first known example of the lightly-built bipedal animal with a beak body plan, aka, the “ostrich” body plan, even though this iteration - the first iteration - had nothing resembling feathers or efficient breathing or hollow bones. In fact, it probably would have breathed primarily based on abdominal muscles, based on its close relatives. And it did have hollow bone walls, much like dinosaurs. Effigia also had a somewhat endothermic body temp - ie, it was closer to warm-bloodedness than modern crocodilians, and may have even been warm-blooded outright. It had five fingers on each hand, though only three of each would have claws; it had four main toes on each foot, unlike the three in theropods, and a little toe raised up (kind of like the fourth toe in theropods). It was rather front heavy, unlike theropods that lean more towards the hip, giving it a forward-leaning appearance.
Diet: Probably herbivorous. While the exact diet of Effigia is murky, the beak of this species indicates it probably would have fed on a variety of plant material, like the later mimics such as Limusaurus and Ornithomimus. However, omnivory was certainly not out of the question.
Behavior: Because Effigia was front-titled, it’s actually not clear whether or not it would have behaved similarly to the animals it resembles. In fact, it doesn’t seem very well adapted for fast movement at all - it looks exactly like the sort of creature that may have tripped over itself on a regular basis. That said, it is entirely possible that it would have balanced itself differently, or adjusted its position in such a way to make up for this oddity in posture. That said, it also had fairly short legs compared to its overall body length, so it’s doubtful that it would have been a fast mover in any position. As such, it probably would have moved slowly throughout its ecosystem, grazing on plants and following fresh vegetation where it came up and utilizing its long neck to reach into areas where food was less accessible, and grabbing on to it for feeding. It may not have been a particularly social animal though, like living archosaurs, it probably would have taken care of its young in some fashion.
Ecosystem: Ghost Ranch was a large floodplain, not quite as forested as the environment had been in earlier times (when they were literally called the “petrified forest”), however, there were still extensive dry forests that experienced dramatic dry and wet seasons each year. These seasons were interspersed with regular flooding, which lead to rapid preservation of a very diverse Late Triassic ecosystem. This was an extremely diverse habitat, with a variety of other reptiles that lived alongside Effigia. There was the slender dinosaur Coelophysis, the weirdly-toothed dinosaur Daemonosaurus, the Silesaurids Kawanasaurus and Eucoelophysis, the Lagerpetid Dromomeron, the early Crocodylomorph Hesperosuchus, the Aetosaur Stenomyti, the phytosaur Redondasaurus, the Drepanosaurs Avicranium and Drepanosaurus, the aquatic archosauriform Vancleavea, the sphenodont Whitakersaurus; coelocanths, ray-finned fish, mystery fish, and even invertebrates such as branchiopods and ostracods. It’s possible that Effigia lived alongside other animals as well, but more research is needed into the exact environment of the Chinle Formation where Effigia was found before that can be confirmed. It is entirely possible that Effigia would have been preyed upon by Coelophysis.
Other: Effigia is so freaking weird, you guys. Like, the Triassic may as well be called the Period in Which Reptiles Tried Out All The Things Dinosaurs Would Later Do (But Only Dinosaurs Would Get Paid For It). It is so similar to later Ornithomimosaurs that at least a few paleontologists used to think that the remains later called Effigia were actually dinosaurs, before their proper reassignment into the Pseudosuchians. This just emphasizes how much different reptiles were trying out new designs and new ideas in the Triassic Period, some of which superficially resembled later dinosaurs - but with surprise twists. It also demonstrates exactly how much crocodile-relatives were diversifying extensively in the Triassic, and how hard they would be hit by the end-Triassic extinction.
~ By Meig Dickson
Sources Under the Cut
Colbert, E. H.. 1947. The little dinosaurs of Ghost Ranch. Natural History 59(9):392-399-427-428.
Heckert, A. B., S. G. Lucas, L. F. Rinehart and A. P. Hunt. 2008. A new genus and species of sphenodontian from the Ghost Ranch Coelophysis Quarry (Upper Triassic: Apachean), Rock Point Formation, New Mexico, USA. Palaeontology 51(4):827-845.
Hunt, A. P., and A. G. Lucas. 1989. Late Triassic vertebrate localities in New Mexico. In S. G. Lucas and A. P. Hunt (eds.), Dawn of the Age of Dinosaurs in the American Southwest, New Mexico Museum of Natural History, Albuquerque 72-101.
Hunt, A. P., and S. G. Lucas. 1993. A new phytosaur (Reptilia: Archosauria) genus from the uppermost Triassic of the western United States and its biochonological significance. In S. G. Lucas and M. Morales (eds.), The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin 3:193-196.
Lucas, S. G., J. A. Spielmann, A. P. Hunt. 2007. Taxonomy of Shuvosaurus, a Late Triassic archosaur from the Chinle Group, American Southwest. New Mexico Museum of Natural History and Science Bulletin 41: 259 - 261.
Lucas, S. G., J. A. Spielmann, and L. F. Rinehart. 2013. Juvenile skull of the phytosaur Redondasaurus from the Upper Triassic of New Mexico, and phytosaur ontogeny. New Mexico Museum of Natural History and Science Bulletin 61:389-400.
Martz, J. W., B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7: e7551.
Nesbitt, S., M. A. Norell. 2006. Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society B: Biological Sciences 273 (1590): 1045-1048.
Nesbitt, S. 2007. The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History 302: 84.
Nesbitt, S. J., M. R. Stocker, B. J. Small and A. Downs. 2009. The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes). Zoological Journal of the Linnean Society 157:814-864.
Nesbitt, S. 2011. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades. Bulletin of the American Museum of Natural History 352: 1 - 292.
Pritchard, A. C., and S. J. Nesbitt. 2017. A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Royal Society Open Science 4:170499.
Renesto, S., J. A. Spielmann, S. G. Lucas and G. T. Spagnoli. 2010. The taxonomy and paleobiology of the Late Triassic (Carnian-Norian: Adamanian-Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha). New Mexico Museum of Natural History and Science, Bulletin 46:1-81.
Rinehart, L. F., S. G. Lucas, A. B. Heckert, J. A. Spielmann, and M. D. Celeskey. 2009. The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. New Mexico Museum of Natural History and Science Bulletin 45:1-260.
Schachner, E. R., P. L. Manning, P. Dodson. 2011. Pelvic and hindlimb myology of the basal archosaur Poposaurus gracilis (archosauria: Poposauroidea). Journal of MOrphology 272 (12): 1464 - 1491.
Schaeffer, B. 1967. Late Triassic fishes from the western United States. Bulletin of the American Museum of Natural History 135(6):285-342.
Sues, H.-D., S. J. Nesbitt, D. S. Berman and A. C. Henrici. 2011. A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B 278:3459-3464.
#Effigia#Effigia okeeffeae#Poposaur#Pseudosuchian#Triassic#Archosaur#Reptile#Triassic Madness#Palaeoblr#Triassic March Madness#Prehistoric Life#Paleontology#Prehistory
198 notes
·
View notes
Text
Daemonosaurus chauliodus

By José Carlos Cortés
Etymology: Demon Reptile
First Described By: Sues et al., 2011
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Theropoda
Status: Extinct
Time and Place: Between 205 and 202 million years ago, in the Rhaetian of the Late Triassic
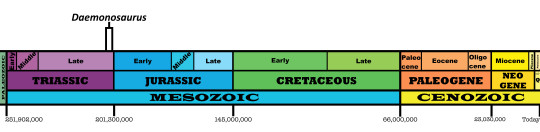
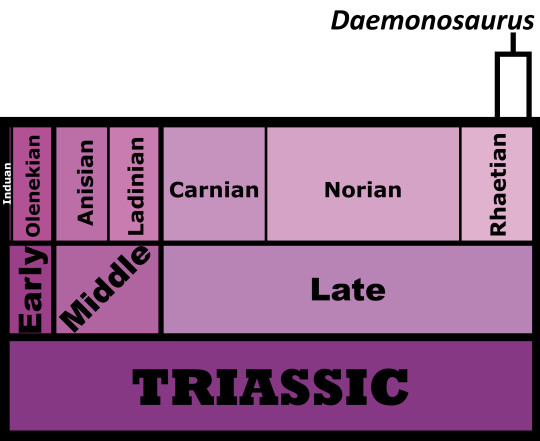
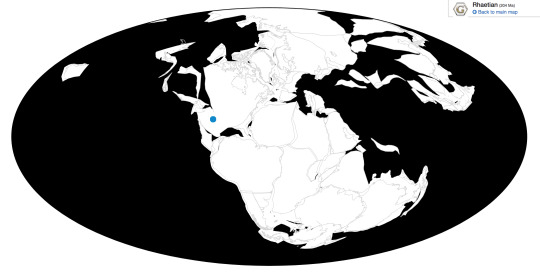
Daemonosaurus is known from the Siltstone Member of the Chinle Formation in New Mexico
Physical Description: Daemonosaurus is an interestingly little dinosaur, and an enigmatic one, with its placement within the dinosaur family tree holding significant importance for how dinosaurs organize themselves in their initial diversification. This was a small, bipedal dinosaur, only about 1.5 meters long and weighing no more than 22 kilograms. While its body was relatively normal of a dinosaur at that time - longer legs than arms, arms built for grasping, long tail and stout torso - its head was downright bizarre. The skull was very short and box-like, rather than long and narrow or long and rectangular like other predatory dinosaurs of the time. Daemonosaurus also featured very long and large teeth in the upper jaw, and teeth that projected forward out of the mouth from both jaws - in short, it looked fairly buck-toothed. It also had a slight notch in its jaws, which could have been used to grab hold of struggling food and trap it there. It also might be an example of paedomorphy - while the head seems fairly juvenile (including rather big eyes), the rest of the body has the fused bones of an adult. As a small dinosaur, Daemonosaurus would have been covered in fluffy protofeathers for thermoregulation.
Diet: Daemonosaurus would have primarily eaten small animals like early mammals and smaller reptiles, though baby dinosaurs from other species wouldn’t have been off the menu.

By Michael B. H., CC BY-SA 3.0
Behavior: Daemonosaurus, being a smaller theropod, wouldn’t have been a very bold predator. Instead, it probably waited in the shadows and undergrowth a lot, looking for a moment to strike at its prey. Hiding in the bushes, it was able to stay safe from larger predators, which frequented the environment. It probably would use its hands and mouth to grab struggling prey, and also defend itself from danger. With its very large eyes, it’s even possible it was somewhat nocturnal, and did most of its hunting in the safety of night. It probably took care of its young, though of course without fossil evidence of such it is hard to tell; and it would have been a fairly active, intelligent animal in its habitat.
Ecosystem: Daemonosaurus lived in the Siltstone Environment of the Chinle Formation, a famous ecosystem showcasing the rise of dinosaurs within North America at the end of the Triassic Period. Daemonosaurus is known from one of the later ecosystems of that formation, which makes its position as a fairly basal theropod somewhat surprising. This was a seasonally arid floodplain, with trivers that would occasionally flood and alternate between that and drying up completely. As such, the plantlife around the floodplain was mostly hardy ferns, ginkgoes, horsetails, and cycads - as well as a fairly dense forest of pine and other coniferous trees. Here Daemonosaurus shared its environment with many other Triassic weirdos, such as the Drepanosaur (lizard-monkey thing) Avicranium, ray-finned fish such as Hemicalypterus and Lophionotus, the phytosaur Redondasaurus, the small Aetosaur Stenomyti; and other Dinosauromorphs. There was the Silesaurid Eucoelophysis, the Lagerpetid Dromomeron, and another theropod dinosaur, Ceolophysis.

By Scott Reid
Other: Is Daemonosaurus a theropod? Probably. But its head is so weird - and its whole body, too - that the position of Daemonosaurus is a question within the early diversification of dinosaur group. Indeed, Daemonosaurus often changes how a phylogenetic tree turns out. This weirdness is something to keep an eye on for now, because the jury is still out - though basal theropod seems likely. And what a weird theropod lineage it represents!
~ By Meig Dickson
Sources under the Cut
Barta, D. E., S. J. Nesbitt, M. A. Norell. 2018. The evolution of the manus of early theropod dinosaurs is characterized by high inter- and intraspecific variation. Journal of Anatomy 232: 80 - 104.
Ezcurra, M.D. (2006). "A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA." Geodiversitas, 28(4):649-684.
Griffin, C. T. 2019. Large neotheropods from the Upper Triassic of North America and the early evolution of large theropod body sizes. Journal of Paleontology: 1 - 21.
Irmis, R. B. 2005. The vertebrate fauna of the Upper Triassic Chinle Formation in northern Arizona. p. 63-88. in S.J. Nesbitt, W.G. Parker, and R.B. Irmis (eds.) 2005. Guidebook to the Triassic formations of the Colorado Plateau in northern Arizona: Geology, Paleontology, and History. Mesa Southwest Museum Bulletin 9.
Nesbitt, S.J., Irmis, R.B., and Parker, W.G. (2005). "A critical review of the Triassic North American dinosaur record." In Kellner, A.W.A., Henriques, D.D.R., & Rodrigues, T. (eds.), II Congresso Latino-Americano de Paleontologia de Vertebrados, Boletim de Resumos. Rio de Janeiro: Museum Nacional/UFRJ, 139.
Sues, H.-D., S. J. Nesbitt, D. S. Berman, A. C. Henrici. 2011. A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B 278 (1723): 3459 - 3464.
Sullivan, C., X. Xu. 2016. Morphological Diversity and Evolution of the Jugal in Dinosaurs. The Anatomical Record 300 (1): 30 - 48.
Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. 861 pp. ISBN 0-520-24209-2.
#Daemonosaurus chauliodus#Daemonosaurus#Dinosaur#Theropod#Prehistoric Life#Paleontology#Palaeoblr#Factfile#Prehistory#Carnivore#Triassic#North America#Theropod Thursday#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
318 notes
·
View notes