#Journal of Systematic Paleontology
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
Average visit duration of Tumblr.com is 10 mins and 25 secs.
Text









#dinosaurs#dinosaur fossils#Comptonatus chasei#Isle of Wight#England#University of Portsmouth#prehistoric creature#fossilized footprints#Journal of Systematic Paleontology#Nick Chase#excavation site#Mary Anning#paleontology#Compton Bay#dinosaur species#skeleton
16 notes
·
View notes
Text

Isisaurus colberti was a sauropod dinosaur that lived in what is now India and Pakistan at the end of the Cretaceous Period, about 70-66 million years ago.
It was part of the titanosaur group of sauropods and had some unusual proportions* compared to its close relatives, with elongated forelimbs and a relatively short chunky neck. Since it's only known from a partial skeleton its full size is unknown – estimates have been made as large as 18m long (~60') but it was probably somewhat smaller, closer to around 11m in length (36').
(*Measurement errors in the original paper resulted in some very weird proportions, but more recent and rigorous reconstructions have made Isisaurus not quite so cursed-looking.)
Like most other titanosaurs it probably lacked the thumb claws seen in other sauropods, and it may also have had some bony osteoderm armor studding its skin.
Coprolites that may represent Isisaurus' poop show evidence of several different fungi that grow on tree leaves in humid tropical and subtropical climates, suggesting that this sauropod was a selective browser somewhat like modern giraffes.
———
NixIllustration.com | Tumblr | Patreon
References:
Jain, Sohan L., and Saswati Bandyopadhyay. "New titanosaurid (Dinosauria: sauropoda) from the Late Cretaceous of central India." Journal of Vertebrate Paleontology 17.1 (1997): 114-136. https://doi.org/10.1080/02724634.1997.10010958
Naish, Darren. "The hands of sauropods: horseshoes, spiky columns, stumps and banana shapes." Tetrapod Zoology, 29 Oct. 2008, https://web.archive.org/web/20121022121526/http://scienceblogs.com/tetrapodzoology/2008/10/29/the-hands-of-sauropods/
Sharma, Neeta, et al. "Fungi in dinosaurian (Isisaurus) coprolites from the Lameta Formation (Maastrichtian) and its reflection on food habit and environment." Micropaleontology 51.1 (2005): 73-82. https://doi.org/10.2113/51.1.73
Wedel, Matt. "How shortweird was Isisaurus?" Sauropod Vertebra Picture of the Week, Aug. 13 2009, https://svpow.com/2009/08/13/how-shortweird-was-isisaurus/
Wikipedia contributors. “Isisaurus” Wikipedia, 27 Dec. 2024, https://en.wikipedia.org/wiki/Isisaurus
Wilson, Jeffrey A., and Paul Upchurch. "A revision of Titanosaurus Lydekker (Dinosauria‐Sauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution." Journal of Systematic Palaeontology 1.3 (2003): 125-160. https://doi.org/10.1017/S1477201903001044
#science illustration#paleontology#paleoart#palaeoblr#isisaurus#titanosaur#sauropod#sauropodomorpha#saurischian#dinosaur#art
305 notes
·
View notes
Text

This ancient crocodile looked kind of like a dolphin
Enalioetes schroederi’s ‘radically different’ body was scaleless, speedy, and lost to researchers for decades.
A set of 135 million-year-old fossils, once thought lost during World War II, are now accounted for and analyzed. According to a study published July 18 in the Journal of Systematic Paleontology, a team of international researchers from Germany and the UK has determined the “well preserved” skull and first neck vertebrae are from what they now call Enalioetes schroederi. But while paleontologists now know they belong to a newly discovered species of ancient marine crocodile, these creatures likely looked more like dolphins than reptiles...
Read more:
https://www.popsci.com/science/ancient-crocodile-dolphin/
382 notes
·
View notes
Text
The trilobite crest of the Heterodynes really raises questions about the history of palaeontology in the setting.
Like, honestly, considering this is a world where autism gives you deeply unpleasant cognitive superpowers that are nevertheless constrained (to some extent) by your background knowledge, it would kind of make sense if a lot of the 'making lists' sorts of natural science were millennia ahead of our timeline. Because that's a science you can do without an industrial revolution behind you, right?
According to wikipedia the trilobite (wildly common as fossils go) first appears in the scientific literature in 1698 when
Rev. Edward Lhwyd published in 1698 in The Philosophical Transactions of the Royal Society, the oldest scientific journal in the English language, part of his letter "Concerning Several Regularly Figured Stones Lately Found by Him", that was accompanied by a page of etchings of fossils.[118] One of his etchings depicted a trilobite he found near Llandeilo, probably on the grounds of Lord Dynefor's castle, he described as "the skeleton of some flat Fish".
A family with the antiquity and inflexibility of the Heterodynes is not going to have adopted their characteristic logo within the past 200 years. 200 years ago is like. Euphrosinia's day. They were already infamous on a massive scale. There is no way.
So does this suggest a classical discipline of palaeontology? Like. To do really useful paleontology you need a fairly systematic process, and I feel like if the scientific method had recognizably developed by the classical period this would have ended the 'wizard spark' era sooner than the Renaissance.
Or did the trilobite just not perish in the Permian mass extinction event and they still exist???
Also oh hey check it out, trilobites are apparently technically a Welsh discovery huh.
#hoc est meum#it's also a setting with time travel#but not like on an ~everyday basis#so i don't know if i want to blame the trilobite on that#tho ig it would be funny if lucrecia somehow caused the heterodyne trilobite by messing with the timeline#the queens do thoroughly upend most assumptions about history too ig#but i still want to poke at it#girl genius#trilobite
98 notes
·
View notes
Text
Fwd: Postdoc: Okinawa_IST.EvolutionaryIchthyology
Begin forwarded message: > From: [email protected] > Subject: Postdoc: Okinawa_IST.EvolutionaryIchthyology > Date: 2 March 2024 at 05:26:52 GMT > To: [email protected] > > > > Okinawa Institute of Science and Technology > Postdoctoral Scholar or Staff Scientist in Ichthyology > > A postdoctoral scholar (PhD 5 years) or staff scientist position > (PhD 5 years) is available in the Macroevolution Unit at OIST. We > seek a highly motivated, expert researcher with a deep knowledge > of fish biodiversity, including but not limited to systematics, > taxonomy, ecology., and the fossil record. The researcher will pursue > groundbreaking projects on ichthyological topics, such as the origins of > the Indo-Pacific biodiversity hotspot, the origins and diversification of > fish groups, description and characterization of species and communities, > and/or the influence of environmental and ecological factors on fish > biodiversity. Exact projects will be developed through discussions with > PI Sallan based on the skills, interests, and ideas of the selected > researcher. > > Responsibilities: > > Develop and lead innovative projects related to the ichthyological > interests of the Macroevolution Unit. > Publish results in high-quality journals. > Present at national and international conferences. > > Qualifications: > > (Required) > > PhD in Ecology and Evolutionary Biology, Marine Biology, Paleontology, > Organismal Biology or related field with dissertation focused on fishes. > Experience and interest in fish biodiversity. > Willingness to develop knowledge about areas and clades of interest to > the unit if outside of past experience (e.g. fishes). > Willingness to learn new methods as needed. > Willingness to travel to museum collections or collect specimens as needed. > Willingness to collaborate with other relevant labs at OIST and outside. > Good spoken and written English skills, including for scientific terms > and concepts. > > (Preferred) > > Experience in either fish morphology, fish phylogenetics, marine field > work and collections, or work with live fishes, depending on area of > expertise. > > Start Date: > Negotiable from April 2024 (if in Japan already) through > mid-2025. Promising senior graduate students and postdocs with time > remaining in their current positions are encouraged to apply. > > Term & Working hours: > Full-time. A postdoctoral position (PhD5 years) is initially for one > year and can be renewed to 2 more years based on performance and mutual > agreement. A staff scientist position (PhD5 years) is initially for > one year and can be extended further based on performance and mutual > agreement. > > Working hours: > > Discretionary working hours > > Compensation & Benefits: > Compensation in accordance with the OIST Employee Compensation Regulations > > Benefits: > > Relocation, housing and commuting allowances > Annual paid leave and summer holidays > Health insurance (Private School Mutual Aid > https://ift.tt/dtH8DFp ) > Welfare pension insurance (kousei-nenkin) > Worker’s accident compensation insurance (roudousha-saigai-hoshou-hoken) > > Application Due Date: > > Applications will continue to be screened until the position is filled. > > Submission Documents: > > Cover letter including statement of research interests > > CV including the publication list > > Contact information for 3 recommendation letters (will be contacted > after first pass) > > Apply At: > > https://ift.tt/Lo6lxkJ > > Lauren Sallan
0 notes
Text
Nueva especie de dinosaurio blindado encontrada en la Isla británica de Wight
Con un cuerpo totalmente acorazado y una dieta estrictamente herbívora, así han descrito a esta nueva especie de dinosaurio recién descrito en la formación Wessex en la Isla británica de Wight. En el estudio publicado en la revista de fomento científico Journal of Systematic Paleontology, los paleontólogos describen la nueva especie como Vectipelta barretti, un dinosaurio acorazado perteneciente…
View On WordPress
#anquilosaurios#Cretácico Temprano#Isla de Wight#Museo de Historia Natural#nueva especie de dinosaurio acorazado#Reinó Unido#Vectipelta barretti
0 notes
Text
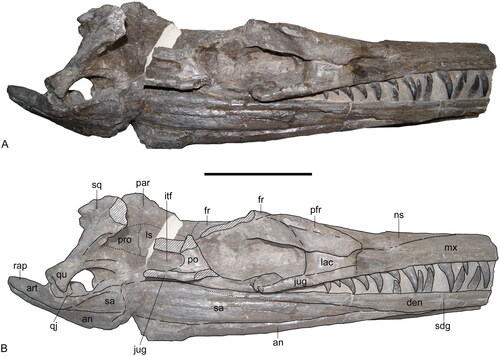
Meemannavis ductrix O’Connor et al., 2021 (new genus and species)

(Type specimen of Meemannavis ductrix [inset in A shows lower jaws in cross section, visualized through CT scanning], from O’Connor et al., 2021)
Meaning of name: Meemannavis = [Chinese paleontologist] Chang Meemann’s bird; ductrix = leader [referring to Chang’s role as the first woman to serve as director of the Institute of Vertebrate Paleontology and Paleoanthropology at the Chinese Academy of Sciences]
Age: Early Cretaceous (Aptian)
Where found: Xiagou Formation, Gansu, China
How much is known: Partial skull and several neck and back vertebrae of one individual.
Notes: Meemannavis was a euornithean, a group including modern birds and some of their closest extinct relatives. It can be distinguished from other euornitheans with preserved skull material from the same locality by having a toothless lower jaw. The tip of its upper jaw was also toothless, but possible tooth sockets are visible further back in the upper jaw of the type specimen.
Although Meemannavis is clearly distinct from other Xiagou Formation euornitheans for which skull material is known, overlapping material has not been found for three previously-named euornitheans from the same locality, Changmaornis, Jiuquanornis, and Yumenornis. It is therefore conceivable that Meemannavis might end up being representing the skull of one of these genera.
Reference: O'Connor, J.K., T.A. Stidham, J.D. Harris, M.C. Lamanna, A.M. Bailleul, H. Hu, M. Wang, and H. You. 2021. Avian skulls represent a diverse ornithuromorph fauna from the Lower Cretaceous Xiagou Formation, Gansu Province, China. Journal of Systematics and Evolution advance online publication. doi: 10.1111/jse.12823
34 notes
·
View notes
Text
Odontochelys semitestacea

By @alphynix
Etymology: Toothed Turtle
First Described By: Lie et al., 2008
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha?, Archelosauria, Pantestudines, Odontochelyidae
Time and Place: Around 232 million years ago, in the Carnian age of the Late Triassic
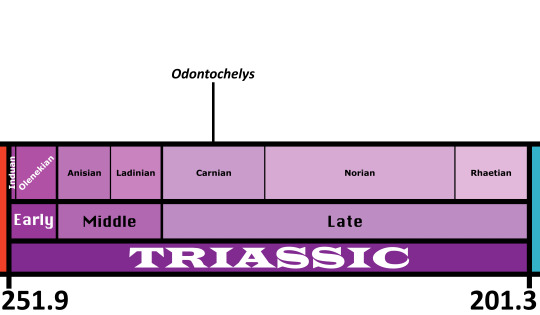
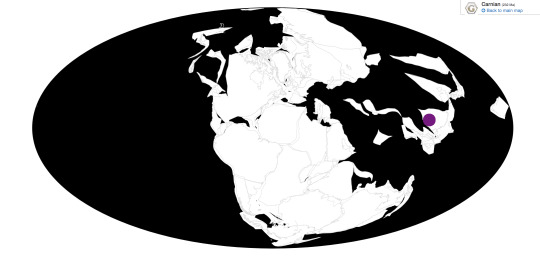
Odontochelys is known from the Lower Member of the Xiaowa Formation of China, commonly known as the Guanling Fauna
Physical Description: Odontochelys is one of the earliest known turtles - preceded by one, possibly two, precursors other than stem members of the family group - and it showcases how this extremely unique group managed to evolve in the chaos that was the Triassic Explosion. It was simultaneously similar to and very different from living turtles, a true transitional organism. Like other reptiles, it had teeth embedded in its jaws, rather than the toothless beak found in turtle mouths. Like turtles, it had the lower plastron extending from its ribs, but unlike living turtles it had no upper shell - instead, it just had widened ribs and no bony shell around its body. The ribs and the vertebrae were put together differently from modern turtles as well, and its skull was more stretched out compared to its living relatives. It didn’t have fused tail bones, and its scapulae were very different from living turtles. It had short limbs and long, thick fingers, as well as a decently sized shell. It was about forty centimeters long from snout to tail tip.
Diet: The diet of Odontochelys is fairly uncertain, despite us having its teeth; though they are small and peg like, we can’t really extrapolate a function since we don’t actually know its precise ecology! They may have been used for stripping plants, but it’s also possible they were used to chipp up algae and other aquatic water plants, or even invertebrates! So, more research there is clearly needed.
Behavior: The life history of Odontochelys is actually a big mystery. It was found in a marine environment, leading initial studies to indicate it was marine. However, it had the hands of a fresh water organism, including fresh water turtles today. Furthermore, studies of other early turtles indicate that turtles first arose on the land, rather than in the water, and later groups would adapt to water life; the limbs of Odontochelys share similarities with tortoises and support a terrestrial lifestyle. So, the ecology of Odontochelys has been a constant battle. That said, there is some evidence that it was actually marine - and may represent an early experiment in ocean life by turtles. One fossil of Odontochelys indicates that it had completely messed up shoulder bones, likely due to a problem in life rather than destruction of the fossil. This pattern resembles decompression sickness, aka the bends, aka the condition caused by a diving animal coming up much too fast from a lower depth. Modern turtles have complex behavioral adaptations to avoid the bends, so Odontochelys may be an early experiment in marine life in a group mostly adapted for terrestrial life. In this transition to ocean life, it not only lacked better physical adaptations for the ocean, but also better behavioral ones, and was stricken with the bends on its trip back to the surface. So, as we try to determine its ecology and behavior, these clued paint a rich tapestry of the world’s most transitional turtle. A pioneer!
Ecosystem: Odontochelys was found in - and thus, the null hypothesis is that it lived in - an ocean environment near the coast of the Tethys sea. This was a deep, open ocean - pelagic, hence the bends and problems Odontochelys faced trying to deal with the ocean. It was a very fertile ecosystem as well, with a variety of Triassic marine animals showcasing the rapid evolution of these groups. Among the invertebrates, there were many different types of Ammonites, plenty of bivalves, brachiopods, and crinoids & sea cucumbers as well. Still, the fascinating part of the ecosystem was the sheer number of marine reptiles. There were Thalattosaurs such as Anshusaurus and Xinpusaurus; Placodonts like Psephochelys and Sinocyamodus; and Ichthyosaurs like Qianicthyosaurus, Guizhouichthyosaurus, Guanlingsaurus, and Callawayia. One of many beautiful deposits of marine animals from this Period - and the many Ichthyosaurs would have been major predators of the relative n00b Odontochelys.
Other: Odontochelys also just looks really weird because it basically looks like your usual turtle except it doesn’t have a freaking shell so here we are with this oddity. It’s transitional in shape, transitional in ecology, transitional in behavior, and just. What the heck. What the heck, Odontochelys. If you need more proof for evolution despite knowing about the dinosaur - bird transition, have I got a friend for you.
~ By Meig Dickson
Sources Under the Cut
Anquetin, J. 2012. Reassessment of the phylogenetic interrelationships of basal turtles (Testudinata). Journal of Systematic Palaeontology 10(1):3-45.
Bradley Shaffer, H., E. McCartney-Melstad, T. J. Near, G. G. Mount, P. Q. Spinks. 2017. Phylogenomic analyses of 539 highly informative loci dates a fully resolved time tree for the major clades of living turtles (Testudines). Molecular Phylogenetics and Evolution 115: 7 - 15.
Feldmann, R. M., C. E. Schweitzer, S. Hu, J. Huang, Q. Zhang, C. Zhou, W. Wen, T. Xie, E. P. Maguire. 2017. A new Middle Triassic (Anisian) cyclidan crustacean from the Luoping Biota, Yunnan Province, China: morphologic and phylogenetic insights. Journal of Crustacean Biology 37 (4): 406 - 412.
Gilbert, S. F. 2007. How the turtle gets its shell. Biology of Turtles: The Structures to Strategies of Life.
Heiss, E. 2010. Functionality and plasticity of turtle-feeding with special emphasis on oropharyngeal structures. Universitat Wien Doctoral Dissertation.
Hess, H., W. Etter, and H. Hagdorn. 2016. Roveacrinida (Crinoidea) from Late Triassic (early Carnian) black shales of Southwest China. Swiss Journal of Paleontology 135(2):249-274.
Joyce, W. G. 2015. The origin of turtles: A paleontological perspective. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 324 (3): 181 - 193.
Lee, M. S. Y. 2013. Turtle origins: Insights from phylogenetic retrofitting and molecular scaffolds. Journal of Evolutionary Biology 26 (12): 2729 - 2738.
Lemell, P., N. Natchev, C. J. Beisser, E. Heiss. 2019. Feeding in Turtles: Understanding Terrestrial and Aquatic Feeding in a Diverse but Monophyletic Group. Feeding in Vertebrates: 611 - 642.
Li, C., and O. Rieppel. 2002. A new cyamodontid placodont from Triassic of Guizhou, China. Chinese Science Bulletin 47(5):403-407.
Li, C., X. C. Wu, O. Rieppel, L. T. Wang, and L. J. Zhao. 2008. An ancestral turtle from the Late Triassic of southwestern China. Nature 456:497-501.
Lu, H., D.-Y. Jiang, R. Motani, P.-G. Ni, Z.-Y. Sun, A. Tintori, S.-Z. Xiao, M. Zhou, C. Ji, W.-L. Fu. 2018. Middle Triassic Xingyi Fauna: Showing turnover of marine reptiles from coastal to oceanic environments. Palaeoworld 27 (1): 107 - 116.
Luo, M., Y.-M. Gong, G. R. Shi, Z.-Q. Chen, J. Huang, S. Hu, X. Feng, Q. Zhang, C. Zhou, W. Wen. 2018. Palaeoecological Analysis of Trace Fossil Sinusichnus sinuosus from the Middle Triassic Guanling Formationin Southwestern China. Journal of Earth Science 29: 854 - 863.
Meredith, R. W., J. Gatesy, M. S. Springer. Molecular decay of enamel matrix protein genes in turtles and other edentulous amniotes. BMC Evolutionary Biology 13: 20.
Neenan, J. M., N. Klein, T. M. Scheyer. 2013. European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications 4: 1621.
Nicholls, E. L., C. Wei, and M. Manabe. 2002. New material of Qianichthyosaurus Li, 1999 (Reptilia, Ichthyosauria) from the Late Triassic of southern China, and implications for the distribution of Triassic icthyosaurs. Journal of Vertebrate Paleontology 22(4):759-765.
Reisz, R. R., J. J. Head. 2008. Palaeontology: Turtle origins out to sea. Nature 456 (7221): 450 - 451.
Rothschild, B. M., V. Naples. 2015. Decompression syndrome and diving behavior in Odontochelys, the first turtle. Acta Palaeontologica Polonica 60 (1): 163 - 167.
Schoch, R. R., H.-D. Sues. 2015. A Middle Triassic stem-turtle and the evolution of the turtle body plan. Nature 523 (7562): 584 - 587.
Shang, Q.-H., and C. Li. 2009. On the occurrence of the ichthyosaur Shastasaurus in the Guanling biota (Late Triassic), Guizhou, China. Vertebrata PalAsiatica 47(3):178-193.
Vermeij, G. J., R. Motani. 2017. Land to sea transitions in vertebrates: the dynamics of colonization. Paleobiology 44 (2): 237 - 250.
Wang, X., G. H. Bachmann, H. Hagdorn, P. M. Sanders, G. Cuny, X. Chen, C. Wang, L. Chen, L. Cheng, F. Meng, and G. Xu. 2008. The Late Triassic black shales of the Guanling area, Guizhou province, south-west China: a unique marine reptile and pelagic crinoid fossil lagerstätte. Palaeontology 51(1):27-61.
Wang, X., X. Chen, C. Wang, L. Cheng. 2009. The Triassic Guanling Fossil Group - A Key GeoPark from a barren mountain, Guizhou Province, China. Notebooks on Geology 3: Chapter 2: 11 - 28.
328 notes
·
View notes
Text
Lagerpeton

By Tas
Etymology: Rabbit Reptile
First Described By: Romer, 1971
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Diapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Crurotarsi, Archosauria, Avemetarsalia, Ornithodira, Dinosauromorpha, Lagerpetidae
Referred Species: L. chanarensis
Status: Extinct
Time and Place: About 235 to 234 million years ago, in the Carnian of the Late Triassic
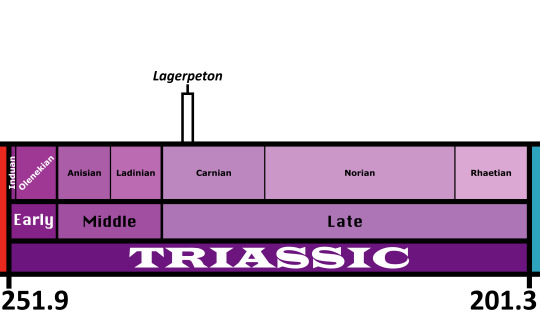
Lagerpeton is known from the Chañares Formation in La Rioja, Argentina

Physical Description: Lagerpeton was named as the Rabbit Reptile, and for good reason - in a lot of ways, it represents a decent attempt by reptiles in trying to do the whole hoppy-hop thing. You might think that it resembles Scleromochlus in that way, and you’d be right! Scleromochlus and Lagerpeton are close cousins, but one is on the line towards Pterosaurs - Scleromochlus - and the other is on the line towards dinosaurs - Lagerpeton. So, hopping around was an early feature that all Ornithodirans (Dinosaurs, Pterosaurs, and those closest to them) shared. Lagerpeton itself was about 70 centimeters in length, with most of that length represented as tail; it was slender and lithe, built for moving quickly through its environment. It had a small head, a long neck, and a thin body. While it had long legs, it also had somewhat long arms, and while it may have been able to walk on all fours it also would have been able to walk on two legs alone. It was digitigrade, walking only on its toes, making it an even faster animal. Its back was angled to help it in hopping and running through its environment, and its small pelvis gave it more force during hip extension while jumping. In addition to all of this, it basically only really rested its weight on two toes - giving it even more hopping ability! As a small early bird-line reptile, it would have been covered in primitive feathers all over its body (protofeathers), though what form they took we do not know.
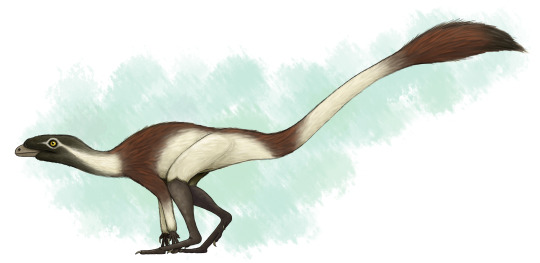
By Scott Reid
Diet: As an early dinosaur relative, it’s more likely than not that Lagerpeton was an omnivore, though this is uncertain as its head and teeth are not known at this time.
Behavior: Lagerpeton would have been a very skittish animal, being so small in an environment of so many kinds of animals - and as such, that hopping and fast movement ability would have aided it in escaping and moving around its environment, avoiding predators and reaching new sources of food (and, potentially, chasing after smaller food itself). Lagerpeton may have also been somewhat social, moving in small groups, potentially families, to escape the predators and chase after prey together, given its common nature in its environment. As an archosaur, Lagerpeton was more likely than not to take care of its young, though we don’t know how or to what extent. The feathers it had would have been primarily thermoregulatory, and as such, they would have helped it maintain a constant body temperature - making it a very active, lithe animal.
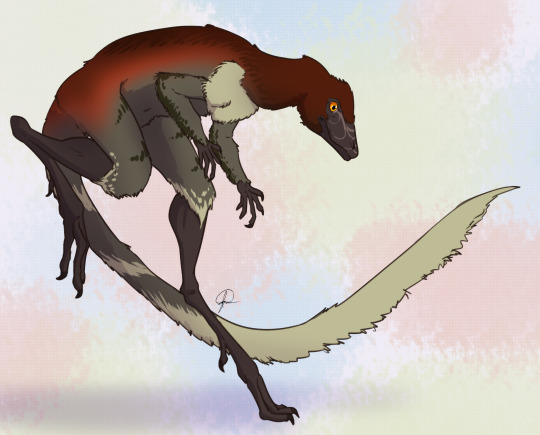
By José Carlos Cortés
Ecosystem: Lagerpeton lived in the Chañares environment, a diverse and fascinating environment coming right after the transition from the Middle to Late Triassic epochs. Given that the first true dinosaurs are probably from the start of the Late Triassic, this makes it a hotbed for understanding the environments that the earliest dinosaurs evolved in. Since Lagerpeton is a close dinosaur relative, this helps contextualize its place within its evolutionary history. This environment was a floodplain, filled with lakes that would regularly flood depending on the season. There were many seed ferns, ferns, conifers, and horsetails. Many different animals lived here with Lagerpeton, including other Dinosauromorphs like the Silesaurid Lewisuchus/Pseudolagosuchus and the Dinosauriform Marasuchus/Lagosuchus. There were crocodilian relatives as well, such as the early suchian Gracilisuchus and the Rauisuchid Luperosuchus. There were also quite a few Proterochampsids, such as Tarjadia, Tropidosuchus, Gualosuchus, and Chanaresuchus. Synapsids also put in a good show, with the Dicynodonts Jachaleria and Dinodontosaurus, as well as Cynodonts like Probainognathus and Chiniquodon, and the herbivorous Massetognathus. Luperosuchus would have definitely been a predator Lagerpeton would have wanted to get away from - fast!

By Ripley Cook
Other: Lagerpeton is one of our earliest derived Dinosauromorphs, showing some of the earliest distinctions the dinosaur-line had compared to other archosaurs. Lagerpeton was already digitigrade - an important feature of Dinosaurs - as shown by its tracks, called Prorotodactylus. These tracks also showcase that dinosaur relatives were around as early as the Early Triassic - and that their evolution, and the rapid diversification of archosauromorphs in general, was a direct result of the end-Permian extinction.
~ By Meig Dickson
Sources Under the Cut
Arcucci, A. 1986. New materials and reinterpretation of Lagerpeton chanarensis Romer (Thecodontia, Lagerpetonidae nov.) from the Middle Triassic of La Rioja, Argentina. Ameghiniana 23 (3-4): 233-242.
Arcucci, A. B. 1987. Un nuevo Lagosuchidae (Thecodontia-Pseudosuchia) de la fauna de los Chañares (Edad Reptil Chañarense, Triasico Medio), La Rioja, Argentina. Ameghiniana 24: 89 - 94.
Arcucci, A., C. A. Mariscano. 1999. A distinctive new archosaur from the Middle Triassic (Los Chañares Formation) of Argentina. Journal of Vertebrate Paleontology 19: 228 - 232.
Bittencourt, J. S., A. B. Arcucci, C. A. Marsicano, M. C. Langer. 2014. Osteology of the Middle Triassic archosaur Lewisuchus admixtus Romer (Chañares Formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs. Journal of Systematic Palaeontology: 1 - 31.
Brusatte, S. L., G. Niedzwiedzki, R. J. Butler. 2011. Footprints pull origin and diversification of dinosaur stem lineage deep into Early Triassic. Proceedings of the Royal Society B: Biological Sciences 278 (1708): 1107 - 1113.
Ezcurra, M. D. 2006. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28(4):649-684.
Ezcurra, M. D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriformes. PeerJ 4: e1778.
Fechner, R. 2009. Morphofunctional evolution of the pelvic girdle and hindlimb of DInosauromorpha on the lineage to Sauropoda (Thesis). Ludwigs Maximillians Universita.
Fiorelli, L. E., S. Rocher, A. G. Martinelli, M. D. Ezcurra, E. Martin Hechenleitner, M. Ezpeleta. 2018. Tetrapod burrows from the Middle-Upper Triassic Chañares Formation (La Rioja, Argentina) and its palaeoecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 496: 85 - 102.
Kammerer, C. F., S. J. Nesbitt, N. H. Shubin. 2012. The first Silesaurid Dinosauriform from the Late Triassic of Morocco. Acta Palaeontological Polonica 57 (2): 277.
Kent, D. V., P. S. Malnis, C. E. Colombi, A. A. Alcober, R. N. Martinez. 2014. Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proceedings of the National Academy of Sciences 111: 7958 - 7963.
Irmis, R. B., S. J. Nesbitt, K. Padian, N. D. Smith, A. H. Turner, D. T. Woody, and A. Downs. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317:358-361.
Jenkins, F. A. 1970. The Chañares (Argentina) Triassic reptile fauna. VII. The postcranial skeleton of the traversodontid Massetognathus pascuali (Therapsida, Cyondontia). Breviora 352: 1 - 28.
Langer, M. C., S. J. Nesbitt, J. S. Bittencourt, R. B. Irmis. 2013. Non-dinosaurian Dinosauromorphs. Geological Society London, Special Publications. 379 (1): 157 - 186.
Marsh, A. D. 2018. A new record of Dromomeron romeri Irmis et al., 2007 (Lagerpetidae) from the Chinle Formation of Arizona, U.S.A. PaleoBios 35:1-8.
Marsicano, C. A., R. B. Irmis, A. C. Mancuso, R. Mundil, F. Chemale. 2016. The precise temporal calibration of dinosaur origins. Proceedings of the National Academy of Sciences of the United States of AMerica 113 (3): 509 - 513.
Martz, J. W., and B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7:e7551:1-71.
Nesbitt, S. J., R. B. Irmis, W. G. Parker, N. D. Smith, A. H. Turner and T. Rowe. 2009. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. Journal of Vertebrate Paleontology 29(2):498-516.
Nesbitt, S. J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 353:1-292.
Perez Loinaze, V. S., E. I. Vera, L. E. Fiorelli, J. B. Desojo. 2018. Palaeobotany and palynology of coprolites from the Late Triassic Chañares Formation of Argentina: implications for vegetation provinces and the diet of dicynodonts. Palaeogeography, Palaeoclimatology, Palaeoecology 502: 31 - 51.
Rogers, R. R., A. B. Arcucci, F. Abdala, P. C. Sereno, C. A. Forster, C. L. May. 2001. Paleoenvironment and taphonomy of the Chañares Formation tetrapod assemblage (Middle Triassic), northwestern Argentina: spectacular preservation in volcanogenic concretions. Palaios 16: 461 - 481.
Romer, A. S. 1966. The Chañares (Argentina) Triassic reptile fauna. I. Introduction. Breviora 247: 1 - 14.
Romer, A. S. 1966. The Chañares (Argentina) Triassic reptile fauna. II. Sketch of the geology of the Rio-Chañares-Rio Gualo Region. Breviora 252: 1 - 20.
Romer, A. S. 1967. The Chañares (Argentina) Triassic reptile fauna. III. Two new gomphodonts, Massetognathus pascuali and M. teruggii. Breviora 264: 1 - 25.
Romer, A. S. 1968. The Chañares (Argentina) Triassic reptile fauna. IV. The dicynodont fauna. Breviora 295: 1 - 25.
Romer, A. S. 1969. The Chañares (Argentina) Triassic reptile fauna. V. A new chiniquodontid cynodont, Probelesodon lewisi - cynodont ancestry. Breviora 333: 1 - 24.
Romer, A. S. 1970. The Chañares (Argentina) Triassic reptile fauna. VI. A chiniquodont cynodont with an incipient squamosal-dentary jaw articulation. Breviora 344: 1 - 18.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. VIII. A fragmentary skull of a large thecodont, Luperosuchus fractus. Breviora 373: 1 - 8.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. IX: The Chanares Formation. Breviora 377: 1 - 8.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. X. Two new but incompletely known long-limbed pseudosuchians. Breviora 378:1-10.
Romer, A. S. 1971. The Chañares (Argentina) Triassic reptile fauna. XI. Two new long-snouted thecodonts, Chanaresuchus and Gualosuchus. Breviora 379: 1 - 22.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XII. The post cranial skeleton of the thecodont Chanaresuchus. Breviora 385: 1 - 21.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XIII. A fragmentary skull of a large thecodont, Luperosuchus fractus. Breviora 389: 1 - 8.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. Lewisuchus admixtus, gen. et sp. Nov., a further thecodont from the Chañares beds. Breviora 390: 1 - 13.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XV. Further remains of the thecodonts Lagerpeton and Lagosuchus. Breviora 394: 1 - 7.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XVI. Thecodont classification. Breviora 395:1-24.
Romer, A. S. 1972. The Chañares (Argentina) Triassic reptile fauna. XVII. The Chañares gomphodonts. Breviora 396: 1 - 9.
Romer, A. S. 1973. The Chañares (Argentina) Triassic reptile fauna. XVIII. Probelesodon minor, a new species of carnivorous cynodont; family Probainognathidae nov. Breviora 401: 1 - 4.
Romer, A. S., and A. D. Lewis. 1973. The Chañares (Argentina) Triassic reptile fauna. XIX. Postcranial materials of the cynodonts Probelesodon and Probainognathus. Breviora 407: 1 - 26.
Romer, A. S. 1973. The Chañares (Argentina) Triassic reptile fauna. XX. Summary. Breviora 413: 1 - 20.
Sereno, P. C., and A. B. Arcucci. 1994. Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. Journal of Vertebrate Paleontology 14(1):53-73.
#Lagerpeton#Dinosaurmorph#Lagerpetid#Triassic#Palaeoblr#Triassic Madness#Ornithodiran#Triassic March Madness#South America#Omnivore#Prehistoric Life#Prehistory#Palaeontology#Lagerpeton chanarensis#dinosaur#paleontology#dinosaurs#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature#factfile
365 notes
·
View notes
Photo




Putting this here because of references & long text. These are WIP. I’ll post the full picture later (Edit: this one link)
I made a Staurikosaurus pricei artwork (paleoart?) because I couldn’t find any good art when I needed it. There are 2 versions of it: scaled and with a speculative integument. Cranium, manus and pes based on Herrerasaurus ischigualastensis. Due to being a rather small animal, with most probably a quite large head, I enlarged the eye opening slighty. Colours inspired by owls, lynx, bobcat and civets because I wanted some nice pattern, in case of forests.
References below, just in case.
Sereno, P. C., & Novas, F. E. (1994). The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, 13(4), 451-476.
Sereno, P. C. (1994). The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, 13(4), 425-450.
Novas, F. E. (1994). New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina. Journal of Vertebrate Paleontology, 13(4), 400-423.
Bittencourt, J. D. S., & Kellner, A. W. A. (2009). The anatomy and phylogenetic position of the Triassic dinosaur Staurikosaurus pricei Colbert, 1970. Zootaxa, 2079(1), e56.
Grillo, O. N., & Azevedo, S. A. (2011). Recovering missing data: estimating position and size of caudal vertebrae in Staurikosaurus pricei Colbert, 1970. Anais da Academia Brasileira de Ciências, 83(1), 61-72. Chicago
Thulborn, T. (2006). On the tracks of the earliest dinosaurs: implications for the hypothesis of dinosaurian monophyly. Alcheringa, 30(2), 273-311.
Scott Hartman’s Herrerasaurus skeleton: https://www.deviantart.com/scotthartman/art/Five-little-piggies-292594542
The Staurikosaurus skeleton featured on Wikipedia: https://en.wikipedia.org/wiki/Staurikosaurus#/media/File:Staurikosaurus_reconstruction.jpg
Saurischian muscle double-check based on RJ Palmer’s T. rex https://www.deviantart.com/arvalis/art/Tyrannosaurus-rex-2018-767137215
And http://www.paleofile.com/Dinosaurs/Theropods/Staurikosaurus.asp (used Google archived thumbnails, as the site was unfortunatelly unavailable when I was collecting references).
#dinosaur#staurikosaurus#herrerasaurus#paleoart#artists on tumblr#digitalart#digital#painting#process#wip
67 notes
·
View notes
Text
Hallan nueva criatura que se extinguió hace miles de años
Hallan nueva criatura que se extinguió hace miles de años
Se trata del Beornus honeyi, una criatura bautizada en homenaje a Beorn debido al aspecto de los molares como inflados. Esta especie es una de las tres nuevas criaturas antiguas de los albores de los mamíferos modernos que han sido descubiertas y descritas en una reciente investigación publicada en el Journal of Systematic Paleontology, en y que sugiere una rápida evolución inmediatamente…

View On WordPress
0 notes
Text

Cadurcodon ardynensis was an odd-toed ungulate that lived in what is now Mongolia during the late Eocene, about 37-34 million years ago.
It was around 2m long (6'6") and, despite its very tapir-like appearance and lack of horns, it was actually closer related to modern rhinoceroses – it was part of a group of early rhino-cousins known as amynodontids, which convergently evolved both hippo-like and tapir-like lifestyles.
Cadurcodon was the most tapir-like of the tapir-like amynodontids, with a short deep skull and retracted nasal bones that indicate it had a well-developed prehensile trunk. Males also had large tusks formed from their upper and lower canine teeth, which may have been used for fighting each other.
———
NixIllustration.com | Tumblr | Patreon
References:
Averianov, Alexander, et al. "A new amynodontid from the Eocene of South China and phylogeny of Amynodontidae (Perissodactyla: Rhinocerotoidea)." Journal of Systematic Palaeontology 15.11 (2017): 927-945. https://doi.org/10.1080/14772019.2016.1256914
Громова, В. [Gromova, V.] Болотные носороги (Amydontidae) Монголии. [Swamp rhinoceroses (Amynodontidae) of Mongolia.] Trudi Paleontol. Inst., Akad. Nauk SSSR 55:85-189 (1954) https://www.geokniga.org/books/13983
Prothero, Donald R., and Robert M. Schoch. Horns, tusks, and flippers: the evolution of hoofed mammals. JHU Press, 2002. http://www.rhinoresourcecenter.com/pdf_files/141/1415340780.pdf
Wall, William P. "Cranial evidence for a proboscis in Cadurcodon and a review of snout structure in the family Amynodontidae (Perissodactyla, Rhinocerotoidea)." Journal of Paleontology (1980): 968-977. https://www.jstor.org/stable/1304363
Wikipedia contributors. “Amynodontidae.” Wikipedia, 17 Dec. 2023, https://en.wikipedia.org/wiki/Amynodontidae
Wikipedia contributors. “Ergilin Dzo Formation.” Wikipedia, 12 Feb. 2024, https://en.wikipedia.org/wiki/Ergilin_Dzo_Formation
#science illustration#paleontology#paleoart#palaeoblr#cadurcodon#amynodontidae#rhinocerotoidea#stem-rhino#perissodactyla#ungulate#mammal#art#he just woke up
275 notes
·
View notes
Photo

Scientists Discover Bug-Eating Reptile That Lived Among Dinosaurs
Delicate fossil reveals a cousin of the modern tuatara
The Morrison Formation is a fossil wonderland. Inside these layers of multi-colored sedimentary stone, the remains of Allosaurus, Stegosaurus, Apatosaurus and more have been found through more than a century of fossil hunting. But these Jurassic rocks contain far more than the bones of “terrible lizards” that lived large. Fossils of small creatures have been filling in what the world of the Morrison Formation was really like, including a new fossil reptile with ties to the modern tuatara.
Named Opisthiamimus gregori, the small creature was a bug-hunting reptile that lived in Wyoming roughly 150 million years ago. Despite the animal’s lizard-like appearance, however, it was not a lizard.
Instead, it belonged to a different group of reptiles known as rhynchocephalians. Described by Smithsonian National Museum of Natural History paleontologist Matthew Carrano and colleagues today in the Journal of Systematic Paleontology, Opisthiamimus was part of that group that flourished in the time of the dinosaurs but is only represented by the tuatara of New Zealand today...
Read more: https://www.smithsonianmag.com/science-nature/scientists-discover-bug-crunching-reptile-that-lived-among-dinosaurs-180980757/
72 notes
·
View notes
Text
Other: Peer Community in Evolutionary Biology
Dear colleagues We invite you to submit your preprints to Peer Community in Evolutionary Biology (PCI Evol Biol, https://evolbiol.peercommunityin.org) in order to obtain a recommendation from this community after peer-reviewing. **Some numbers:** -We're now more than 400 editors at PCI Evol biol -We've received 107 submissions of preprints. We've recommended 46 and are still evaluating 24 of them. -Mean time to 1st decision: 49 days PCI Evol Biol is a group of scientists who review and recommend preprints in evolutionary biology, free of charge and transparently. It is a research community working for researchers to create a new publication system that is completely free of charge and independent of private publishers. This community already includes 400 researchers (the full list can be found at https://evolbiol.peercommunityin.org/public/recommenders) motivated to review and recommend preprints. **Some important points:** -PCI Evol Biol is free for all users (readers AND authors); -PCI Evol Biol is supported by scientific societies including the Society for the Study of Evolution, the Society of Systematic Biologist, the Nordic Society Oikos, the French Ecology and Evolution Society (the SFE2), etc. (https://peercommunityin.org/who-supports-peer-community-in/); -PCI Evol Biol does not publish the preprints (they stay on the preprint server) but the associated reviews and recommendations; -PCI Evol Biol does not evaluate preprints already submitted for publication in a journal, in order to avoid parallel reviewings; -Authors can submit their preprints to traditional journals once they have been recommended by PCI Evol Biol. Reviews obtained before the recommendation can help improve the quality of the preprint; -Journals such as JEB, Evolution, Ecology Letters, Plos Biol, TREE, Evolutionary Applications, etc. (https://peercommunityin.org/pci-friendly-journals/) indicated that they will consider the preprints recommended by PCI Evol Biol and that they will take into account the reviews and recommendations of PCI Evol Biol in their own editorial process if adequate. -PCI Evol Biol's recommendations are transparent (reviews and recommendations are published, recommendations are signed, reviewers can remain anonymous if they wish); -Only reviews leading to a recommendation are published; -A preprint recommended by PCI Evol Biol is scientifically validated and can therefore be cited; -PCI Evol Biol's recommendations have a DOI, are referenced and can therefore also be cited; -Four other communities (PCI Ecology, PCI Paleontology, PCI Animal Science, PCI Entomology) also exist. -Anonymous submissions are possible To submit your preprints, simply go to https://evolbiol.peercommunityin.org click the 'Submit a preprint' button and follow the instructions. We hope to have aroused your interest, The Managing Board of PCI Evol Biol https://evolbiol.peercommunityin.org [email protected] https://youtu.be/4PZhpnc8wwo Thomas Guillemaud
1 note
·
View note
Text
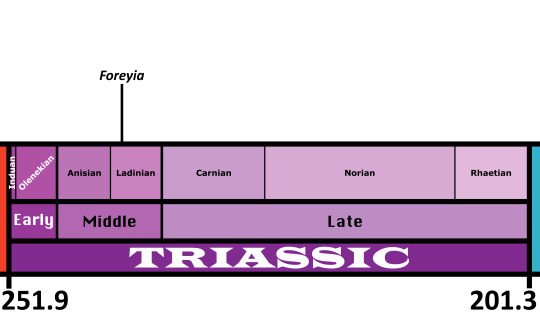
Foreyia maxkuhni

Etymology: Forey’s Fish
First Described By: Cavin et al., 2017
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Actinistia, Coelacanthiformes, Latimerioidei, Latimeriidae,
Time and Place: Foreyia lived about 240.91 million years ago, in the Ladinian of the Middle Triassic
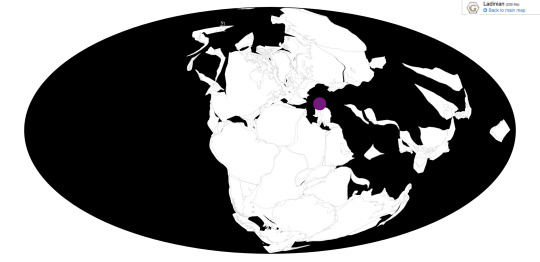
Foreyia is known from the Prosanto Formation of Switzerland
Physical Description: Foreyia was a Coelacanth, a sort of Lobe-Finned Fish once thought to be a unique feature of prehistoric ecologies, and now known to survive in two different species today. This is wild, of course, but Coelacanths were weird in many different ways throughout their prehistoric tenure. Foreyia looked only a little similar to its cousins - it had a huge head compared to other Coelacanths, as well as a beak - something that you really only see in ray-finned fish and tetrapods - an underbite, and a horn on its head. That said, its body looks similar enough to other coelacanths, with dorsal fins and a wide tail fin, and large forelimb fins. However, the large size of its head makes it look like a Coelacanth that had been squished extensively, with the body looking quite short and squished compared to its close relative Ticinepomis. We aren’t sure as to what colors it may have been, but since it lived in a tropical region and may have had a similar niche as a Parrotfish, it is reasonable to suppose it may have been colorful. It was also very small, only around 20 centimeters long.
Diet: Given the beak, it’s likely that Foreyia ate hard food, potentially even similarly to living parrotfish - by chipping off algae covered rocks and reef builders, or potentially eating small shelled animals in the tidal pools.
Behavior: Foreyia’s head was huge, and so that likely impacted its swimming style, though it is unclear how. Modern Coelacanths moving very slowly through the water, moving their fins to help propel through. Since Foreyia reduced the size of its fins to an extent, it’s possible that the tail (which was very large compared to the rest of the body) was more vital in swimming, and the head may have been used to help steer (though this is just a hypothesis). The bony shield may have also been helpful in protecting Foreyia from danger, which makes sense ans many predators were present in its environment. This is at least somewhat supported by its strong clavicle muscles. It probably would have spent a significant amount of time grazing, either on small crunchy invertebrates or algae. Today, Coelacanths don’t seem to shoal much; however, that may not have been the case in the past, so we can’t infer a loner lifestyle for Foreyia.
Ecosystem: Foreyia lived in a coastal shelf environment, specifically in a tidal basin that would often become anoxic as the tide went out during the day, meaning that Foreyia had to move around a lot to avoid suffocation. This environment was probably at least somewhat reef or tidal pool like, with a variety of fish swimming around in the ecosystem. There were many ray-finned fish here, including Peltopleurus, Habrichthys, Archaeosemionotus, Saurichthys, Ctenognathichthys, Besania, Prosanticthys, Eoeugnathus, and Ducanicthys. There was also another Coelacanth, Ticinepornis. As for tetrapods, there was the Pachypleurosaur Neusticosaurus, the Helveticosaurid Eusaurosphargis, and the Tanystropheid Macrocnemus. These reptiles would have been major predators of Foreyia.
Other: Foreyia is so weird in its shape that the original authors proposed it was Heterochronic - ie, different aspects of baby Coelacanth anatomy were retained in order to provide useful adaptations for Foreyia. However, some later researchers have cast doubts on this idea, given that baby Coelacanths don’t… actually seem to resemble Foreyia. Clearly, the jury is still out!
~ By Meig Dickson
Sources Under the Cut
Arratia, G., and A. Herzog. 2007. A New Halecomorph Fish from the Middle Triassic of Switzerland and its Systematic Implications. Journal of Vertebrate Paleontology 27(4):838-849.
Bürgin, T., U. Eichenberger, H. Furrer and K. Tschanz. 1991. Die Prosanto Formation - eine fischreiche Fossil-Lagerstätte in der Mitteltrias der Silvretta-Decke (Kanton Graubünden, Schweiz). Eclogae Geologicae Helvetiae 84:921-990.
Bürgin, T., and A. Herzog. 2002. Die Gattung Ctenognathichthys (Actinopterygii, Perleidformes) aus der Prosanto-Formation (Ladin, Mitteltrias) Graubündens (Schweiz), mit der Beschreibung einer neuen Art, C. hattichi sp. nov. Eclogae Geologicae Helvetiae 95:461-469.
Carroll, R. L., and P. Gaskill. 1985. The nothosaur Pachypleurosaurus and the origin of plesiosaurs. Philosophical Transactions of the Royal Society B 309:343-393.
Cavin, L., H. Furrer, and C. Obrist. 2013. New coelacanth material from the Middle Triassic of eastern Switzerland, and comments on the taxic diversity of actinistans. Swiss Journal of Geosciences 106:161-177.
Cavin, L., B. Mennecart, C. Obrist, L. Costeur, and H. Furrer. 2017. Heterochronic evolution explains novel body shape in a Triassic coelacanth from Switzerland. Scientific Reports 7(13695):1-7.
Ferrante, C., L. Cavin, H. Furrer, R. Martini. 2018. Coelacanths from the Middle Triassic of Switzerland show unusual morphology. Swiss Geoscience Meeting.
Fraser, N., and H. Furrer. 2013. A new species of Macrocnemus from the Middle Triassic of the eastern Swiss Alps. Swiss Journal of Geoscience 106:199-206.
Herzog, A. 2001. Peltoperleidus obristi sp. nov., ein neuer, kleiner Strahlenflosser (Actinopterygii, Perleidiformes) aus der Prosanto-Formation (Mitteltrias) von Graubünden (Schweiz). Eclogae Geologicae Helvetiae 94:495-507.
Herzog, A. 2003. Eine Neubeschreibung der Gattung Eoeugnathus Brough, 1939 (Actinopterygii; Halecomorphi) aus der alpinen Mitteltrais Graubündes (Schweiz). Palaeontologische Zeitschrift 77(1):223-240.
Herzog, A., and T. Bürgin. 2005. A new species of the genus Besania Brough 1939 from the Middle Triassic of Canton Grisons (Switzerland) with a discussion of the phylogenetic status of the taxon. Eclogae Geologicae Helvetiae 98:113-122.
McMenamin, M.A. S. 2018. Coelacanth Vestiges. Deep Time Analysis, Springer Publishing.
Scheyer, T. M., J. M. Neenan, T. Bodogan, H. Furrer, C. Obrist and M. Plamodon. 2017. A new, exceptionally preserved juvenile specimen of Eusaurosphargis dalsassoi (Diapsida) and implications for Mesozoic marine diapsid phylogeny. Scientific Reports 7:4406.
#foreyia#foreyia maxkuhni#coelacanth#Triass March Madness#triassic madness#triassic#prehistoric life#paleontology
298 notes
·
View notes
Text
Silesaurus opolensis

By José Carlos Cortés
Etymology: Silesian Reptile
First Described By: Dzik, 2003
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Silesauridae, Sulcimentisauria
Time and Place: Around 230 million years ago, in the Carnian of the Late Triassic
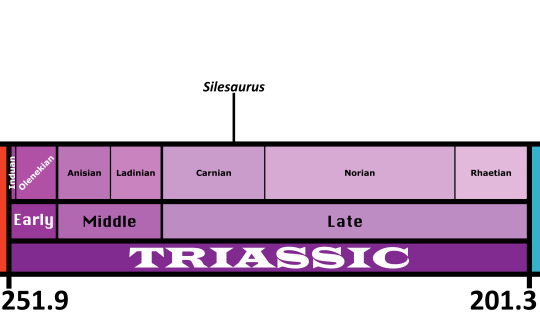
Silesaurus is known from the Drawno Beds Formation of Poland

Physical Description: Silesaurus was a very common almost-dinosaur - specifically, the titular member of the Silesaurids - from Poland in the middle of the Triassic period. It was a quadrupedal animal, about 2.3 meters long and small enough that it most likely would have been covered in fluff. Like other Silesaurids, it was long - with a long neck and a long tail, and long limbs easily reaching to the ground. Like other Silesaurids, it almost had its legs directly underneath its body - it didn’t seem to have an open acetabulum, but rather a partially open one, though Silesaurids are something of a confusing case on this regard. It had a small head, with a little beak in the front, and small sharp conical teeth. In addition, it had very large eyes, giving it distinctly good eyesight, as well as a decent sense of smell. As a quadruped, its front limbs were rather extended, while the hind limbs did a lot of the weight support.
Diet: Interestingly enough, though for a long time it was thought that Silesaurus was an herbivore, recent fossil poop studies have indicated it was an insectivore - similar to living birds.

By Ripley Cook
Behavior: If Silesaurus really was an insectivore, it would have spent much of its time moving around in its environment, examining it closely for insects and pecking at the ground with its small beak for food. This pecking may have even looked a lot like that of living birds - the large eyes probably would have prevented the eyes from moving much, like in living birds, and as such Silesaurus would have had to bob its head in order to see. It would have been very careful in its environment, making sure to look for food and avoid predatory dangers in its environment. It would probably have been social - corroborated by the fact that it was very common in its environment - and taken care of its young to some extent. As an Archosaur, it would have been warm blooded as well, and very active.
Ecosystem: Silesaurus lived in a Mediterranean-esque scrub environment, with summer monsoon seasons and dry winters. The swampy coniferous forest and fernlands it lived in had a wide variety of animals, placing Silesaurus in a unique ecosystem of the Carnian age. This is actually quite important, given that the Carnian Pluvial Event - a minor mass extinction that helped to spur Dinosaurian evolution - occurred around the same time, leading to Silesaurus and its contemporaries being a snapshot of this change. A variety of invertebrates lived alongside this animal, giving it plenty of food, as well as Temnospondyls such as Metoposaurus and Cyclotosaurus, the Aetosaur Stagonolepis, the Phytosaur Parasuchus, the Rauisuchid Polonosuchus, the Sharovpterygid Ozimek, and probably some Tuataras and maybe even a Pterosaur. A small community, but a beautiful portrait of this time period, with its fair share of weirdos and precursors. And Silesaurus would have had a lot to look out for with Polonosuchus at just the right size to attack it.
Other: Do not let Silesaurus’s weird similarities to birds fool you - it wasn’t an early bird, but a showcase of how bird-like traits kept popping up over and over again in the line of animals leading up to them. In fact, the evolution of such a specialized feeding system is notable in such an early member of the Ornithodirans. Silesaurids in general are extremely interesting because of their mixture of characters - similar to dinosaurs, but not quite there; with legs almost directly underneath the body, but other traits that match the later Ornithischians. The problem with Ornithischians - the bird-hipped dinosaurs - is that they have a bit of a ghost lineage, aka even though they should have showed up at the same time as Theropods and Sauropodomorphs, they don’t until the Early Jurassic. This isn’t as big of a gap as some other examples, but it is notable. Silesaurids have come out as Ornithischians just enough times in phylogenetic studies to bring it up as a question - are they almost-dinosaurs? The first bird-hips (without having bird-hips)? Something else entirely? More research is necessarily, but one thing is for sure - this group of weird dinosaurian experiments is one to keep an eye on, especially as we learn more and more about them.
~ By Meig Dickson
Sources Under the Cut
Cau, A. 2018. The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana 57 (1): 1 - 25.
Dzik, J. 2003. A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology 23(3):556-574.
Dzik, J. and T. Sulej. 2007. A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Palaeontologia Polonica 64:3-27.
Ezcurra, M. D. 2006. A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan & Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28(4):649-684.
Griffin, C. T., and S. J. Nesbitt. 2016. The femoral ontogeny and long bone histology of the Middle Triassic (?late Anisian) dinosauriform Asilisaurus kongwe and implications for the growth of early dinosaurs. Journal of Vertebrate Paleontology 36(3):e1111224:1-22.
Irmis, R. B., S. J. Nesbitt, K. Padian, N. D. Smith, A. H. Turner, D. T. Woody, and A. Downs. 2007. A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs. Science 317:358-361.
Jagt, J. W. M., G. Hebda, S. Mitrus, E. Jagt-Yazykov, A. Bodzioch, D. Konietzko-Meier, K. Kardynał, K. Gruntmejer. 2015. Field Guide, Conference Paper, European Association of Vertebrate Palaeontologists XIII Annual Meeting.
Kammerer, C. F., S. J. Nesbitt, and N. H. Shubin. 2012. The first basal dinosauriform (Silesauridae) from the Late Triassic of Morocco. Acta Palaeontologica Polonica 57(2):277-284.
Langer, M. C., M. D. Ezcurra, J. S. Bittencourt and F. E. Novas. 2010. The origin and early evolution of dinosaurs. Biological Reviews 85:55-110.
Martínez, R. N., and O. A. Alcober. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 4(2 (e4397)):1-12.
Martz, J. W., and B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7:e7551:1-71.
Nesbitt, S. J., C. A. Sidor, R. B. Irmis, K. D. Angielczyk, R. M. H. Smith and L. A. Tsuji. 2010. Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature 464:95-98.
Nesbitt, S. J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 353:1-292.
Qvarnström, M., J. V. Wernström, R. Piechowski, M. Tałanda, P. E. Ahiberg, G. Niedźwiedzki. 2019. Beetle-bearing coprolites possibly reveal the diet of a Late Triassic dinosauriform. Royal Society Open Science 6 (3): 181042.
#silesaurus#silesaurus opolensis#dinosauromorph#palaeoblr#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
244 notes
·
View notes